Наращивание ресниц «Лисий эффект» — Artbar Moscow
Администрация сайта https://artbar.moscow/ (далее Сайт) с уважением относится к правам посетителей Сайта. Мы безоговорочно признаем важность конфиденциальности личной информации посетителей нашего Сайта. Данная страница содержит сведения о том, какую информацию мы получаем и собираем, когда Вы пользуетесь Сайтом. Мы надеемся, что эти сведения помогут Вам принимать осознанные решения в отношении предоставляемой нам личной информации. Настоящая Политика конфиденциальности распространяется только на Сайт и на информацию, собираемую этим сайтом и через его посредство. Она не распространяется ни на какие другие сайты и не применима к веб-сайтам третьих лиц, с которых могут делаться ссылки на Сайт.
Сбор информации
Когда Вы посещаете Сайт, мы определяем имя домена Вашего провайдера и страну (например, «https://artbar.
Сайт собирает только личную информацию, которую Вы предоставляете добровольно при посещении или регистрации на Сайте. Понятие «личная информация» включает информацию, которая определяет Вас как конкретное лицо, например, Ваше имя, телефон или адрес электронной почты.
Сайт применяет технологию «cookies» («куки») для создания статистической отчетности. «Куки»
представляет собой небольшой объем данных, отсылаемый веб-сайтом, который браузер Вашего компьютера
сохраняет на жестком диске Вашего же компьютера. В «cookies» содержится информация, которая может
быть необходимой для Сайта, — для сохранения Ваших установок вариантов просмотра и сбора
статистической информации по Сайту, т.е. какие страницы Вы посетили, что было загружено, имя домена
интернет-провайдера и страна посетителя, а также адреса сторонних веб-сайтов, с которых совершен
переход на Сайт и далее. Однако вся эта информация никак не связана с Вами как с личностью.
«Cookies» не записывают Ваш адрес электронной почты и какие-либо личные сведения относительно Вас.
«Куки»
представляет собой небольшой объем данных, отсылаемый веб-сайтом, который браузер Вашего компьютера
сохраняет на жестком диске Вашего же компьютера. В «cookies» содержится информация, которая может
быть необходимой для Сайта, — для сохранения Ваших установок вариантов просмотра и сбора
статистической информации по Сайту, т.е. какие страницы Вы посетили, что было загружено, имя домена
интернет-провайдера и страна посетителя, а также адреса сторонних веб-сайтов, с которых совершен
переход на Сайт и далее. Однако вся эта информация никак не связана с Вами как с личностью.
«Cookies» не записывают Ваш адрес электронной почты и какие-либо личные сведения относительно Вас.
Кроме того, мы используем стандартные журналы учета веб-сервера для подсчета количества посетителей
и оценки технических возможностей нашего Сайта. Мы используем эту информацию для того, чтобы
определить, сколько человек посещает Сайт и организовать страницы наиболее удобным для пользователей
способом, обеспечить соответствие Сайта используемым браузерам, и сделать содержание наших страниц
максимально полезным для наших посетителей. Мы записываем сведения по перемещениям на Сайте, но не
об отдельных посетителях Сайта, так что никакая конкретная информация относительно Вас лично не
будет сохраняться или использоваться Администрацией Сайта без Вашего согласия
Мы используем эту информацию для того, чтобы
определить, сколько человек посещает Сайт и организовать страницы наиболее удобным для пользователей
способом, обеспечить соответствие Сайта используемым браузерам, и сделать содержание наших страниц
максимально полезным для наших посетителей. Мы записываем сведения по перемещениям на Сайте, но не
об отдельных посетителях Сайта, так что никакая конкретная информация относительно Вас лично не
будет сохраняться или использоваться Администрацией Сайта без Вашего согласия
Чтобы просматривать материал без «cookies», Вы можете настроить свой браузер таким образом, чтобы она не принимала «cookies» либо уведомляла Вас об их посылке (различны, поэтому советуем Вам справиться в разделе «Помощь» и выяснить, как изменить установки машины по «cookies»).
Совместное использование информации
Администрация Сайта не продает и не отдает в пользование Вашу личную информацию третьим сторонам. Мы
также не раскрываем предоставленную Вами личную информацию за исключением случаев предусмотренных
законодательством (укажите страну, где расположен сервер).
Мы
также не раскрываем предоставленную Вами личную информацию за исключением случаев предусмотренных
законодательством (укажите страну, где расположен сервер).
Отказ от ответственности Помните, передача информации личного характера при посещении сторонних сайтов, включая сайты компаний-партнеров, даже если веб-сайт содержит ссылку на Сайт или на Сайте есть ссылка на эти веб-сайты, не подпадает под действия данного документа. Администрация Сайта не несет ответственности за действия других веб-сайтов. Процесс сбора и передачи информации личного характера при посещении этих сайтов регламентируется документом «Защита информации личного характера» или аналогичным, расположенном на сайтах этих компаний. Настоящее Соглашение о конфиденциальности и защите персональных данных (далее — Соглашение) регулирует отношения по обработке (сбор, накопление, хранение, уточнение, использование, передачу, блокирование, уничтожение)
Эффекты наращивания ресниц
Эффекты наращивания ресниц
Существующие технологии в
наращивании ресниц, помогают придать взгляду любой женщины привлекательность и
выразительность, делают его особенным и притягательным. Добиться нужного
результата помогут эффекты наращивания ресниц.
Добиться нужного
результата помогут эффекты наращивания ресниц.
Из популярных (классических) эффектов наращивания выделяют: «Натуральный эффект», «Лисий эффект», «Беличий эффект», «Кукольный эффект», «Разреженный эффект» и «Мультицветовой эффект».
Для придания каждым глазам уникального и идеального взгляда в зависимости от формы глаз необходимо смоделировать нужный эффект. Можно воспользоваться схемой классических эффектов либо комбинируя эффекты между собой, изгибы и длинны.
Мы
моделируем, то есть, как бы заново создаем ресницы, такие как нам нужно! Это
понятие пришло в искусство наращивания из визажа, и стало в свою очередь
очень популярным. Моделирование помогает скорректировать форму глаза,
визуально увеличить глаз, «закрыть» нависшее веко.
Кому-то больше подходит легкий изгиб, кому-то
максимально закрученные ресницы, например девушкам с азиатским типом глаз изгиб,
как правило, делают максимальный.
Моделирование помогает выбрать тот эффект ресниц который вам хочется
В первую очередь, рассмотрим глаза нашего клиента. Они
бывают разной формы, величины, выразительности, расположению относительно друг
друга. Условно их можно разделить:
— По форме глазной щели различают миндалевидные или щелевидные глаза
(монгольские, японские).
— По размеру – круглые, большие, маленькие.
— По выразительности – нормально посаженные, глубоко посаженные, или выпуклые.
Если условно провести прямую через внешний и
внутренний уголки глаз – получим, так называемую, ось. Если эта ось
горизонтальная, значит, мы имеем дело с классическим глазом.
Случаи, когда ось, стремится вверх (внешний угол
выше внутреннего), чаще встречается у восточных глаз. Европейцам больше
свойственна ниспадающая ось (наружные уголки ниже внутренних).
Европейцам больше
свойственна ниспадающая ось (наружные уголки ниже внутренних).
Глаза миндалевидной формы принято считать
идеальными. Расстояние между ними равно их длине, при этом внешний внутренние углы лежат на одной прямой.
Задача мастера-лешмейкера, с помощью процедуры наращивания, приблизить глаза к идеальной (миндалевидной) форме. Для этого используются разные по величине изгибы ресниц для наращивания.
Например, для того чтобы получить томный, взгляд выбираем ресницы с легким завитком, это зрительно удлинит разрез глаз. Чтобы сделать взгляд невинным и озорным, выбираем, наоборот, более подкрученные реснички.
Многие клиенты хотят нарастить ресницы, просто для того, чтобы подчеркнуть глазки и не пользоваться косметикой. Для этого есть реснички с небольшим завитком, максимально повторяющим изгиб родных ресниц.
Популярные изгибы ресниц:
Рекомендации по выбору изгиба ресниц для
наращивания:
изгиб B Хорошо подходит тем, у кого форма глаз близка к
идеальной. Он будет повторять изгиб родных ресниц, тем самым подчеркнет
естественный контур глаз. Смотрится очень натурально.
Он будет повторять изгиб родных ресниц, тем самым подчеркнет
естественный контур глаз. Смотрится очень натурально.
— изгиб C Самый популярный. Подходит тем, кто обычно поднимает
ресницы тушью. Зрительно раскрывает глаза.
— изгиб D Этот изгиб распахивает глаза. Подойдет тем, кто
активно подкручивает реснички, а так же у кого опущенное веко или глубокая
глазница, от которой нужно отвлечь внимание.
— изгиб U
(СС) Самый сильный. Предназначен для
тех, кто хочет добиться кукольного эффекта, или пользуется тенями для глаз,
потому что при таком макияже, в особенности нужно подчеркнуть ресницы.Не
подойдет женщинам в возрасте. Категорически НЕ подходит выпуклым (круглым) глазам,
а также в случаях с приподнятым внешним углом.
— изгиб L Создан для глубоко посаженных и азиатских глаз.
Подойдет девушкам, которые носят очки.
— изгиб L+ Более плавный, чем изгиб L. При этом визуально
открывает глаз и приподнимает опущенное веко.
Помимо стремления к идеальному миндалевидному глазу мы также можем создавать разные эффекты взгляда, самые популярные из них:
1. Натуральный эффект —
Также подобную технологию называют классическим наращиванием, что подразумевает под собой создание ресниц с натуральным объемом. Стоит заметить, что все эффекты наращивания ресниц хороши, но результат этого метода является более естественным. Для того, чтобы его выполнить, используются искусственные ресницы одной длины и начинают прикрепляться, начиная с внешнего угла и постепенно следую уже к внутреннему.Для создания натурального эффекта используются ресницы длиной от восьми до десяти миллиметров – это зависит от длины самих натуральных ресниц женщины, ее типа и состояния.
Мы рекомендуем использовать этот метод тем женщинам, которые любят естественную красоту и хотят достигнуть максимально натурального результата.
2. Лисий эффект — Помогает визуально удлинить глаз
в сторону внешнего угла, делает глаз уже, немного опускает верхнее веко,
создает хитрый, томный взгляд. Достигается посредством плавного перехода от
минимальной длины на внутреннем уголке глаза 7-8 мм. до максимальной длины на
внешнем уголке глаза (выбирается клиентом и мастером). Обязательно необходимо
применять если глаза выпуклые или круглые (выбирать не сильные изгибы на
середине глаза), близко посаженые глаза. Также подойдет тем у кого опущенные
уголки (при этом необходимо использовать по всей длине средний или еле
выраженный изгиб, плавно увеличивая изгиб и длину к внешнему углу глаза,
заканчивая последние 5 мм. плавным уменьшение длины и оставляя сильный изгиб)
Достигается посредством плавного перехода от
минимальной длины на внутреннем уголке глаза 7-8 мм. до максимальной длины на
внешнем уголке глаза (выбирается клиентом и мастером). Обязательно необходимо
применять если глаза выпуклые или круглые (выбирать не сильные изгибы на
середине глаза), близко посаженые глаза. Также подойдет тем у кого опущенные
уголки (при этом необходимо использовать по всей длине средний или еле
выраженный изгиб, плавно увеличивая изгиб и длину к внешнему углу глаза,
заканчивая последние 5 мм. плавным уменьшение длины и оставляя сильный изгиб)
3. Беличий эффект — Помогает визуально раскрыть глаз вверх, делая его
более круглым, распахнутым. Достигается посредством перехода к максимальной
длине от середины века, а также постепенным уменьшением длины начиная с 2/3 века
к внешнему углу. Подходит для глубоко посаженых глаз, раскосой форме глаза,
если есть нависшее веко», при опущенных уголках глаз.
4. Кукольный эффект — Еще одним видом эффектов наращивания ресниц является кукольный эффект. Он идеально подходит для молодежной дискотеки, вечеринки или торжественного мероприятия. Девушка с такими ресницами будет выглядеть очень креативно, оригинально и эффектно.
Особенностями является поресничное наращивание и использование очень длинных искусственных ресниц. В результате процедуры ресницы выходят действительно кукольными, потому эффект и носит подобное название.
5. Разреженный эффект — Данный метод, как и натуральный, выглядит максимально естественно, но при этом требует максимально кропотливой работы, чем предыдущие эффекты наращивания ресниц.
Для этого, как и в предыдущем методе, берутся искусственные ресницы разной длины и накладываются в определённом порядке на заранее обговоренном с клиенткой расстоянии.
В итоге, как и было сказано выше, они выглядят максимально естественными, а кроме того еще и густыми.
6. Мультицветовой эффект — В основном, этот метод становится популярным в преддверии какого-либо праздника или мероприятия, однако и в повседневной жизни его можно использовать.
Мультицветовой эффект — В основном, этот метод становится популярным в преддверии какого-либо праздника или мероприятия, однако и в повседневной жизни его можно использовать.
Как вы уже, наверное, догадались, эффект создается с помощью искусственных ресничек разных оттенков. В зависимости от желаний клиентки, используется один или несколько цветов при наращивании. Можно нарастить и несколько цветных ресничек, тогда они будут отлично сочетаться с натуральными черными.
16.10.2016
Перейти к списку новостей
Продолжить
Как правильно выбрать эффект при наращивании ресниц и не разочароваться
Виды и эффекты наращивания ресниц
Эффекты наращивания ресниц: многообразие видов и форм. Процедура наращивания ресничек помогает создать выразительные и распахнутые глаза. В итоге получается притягательный взгляд, который придает дополнительную красоту и шарм своей обладательнице. На первый взгляд кажется, что наращённые ресницы не отличаются между собой. Однако применение различных техник материалов и видов ресничек дает абсолютно разнообразные эффекты. На конечный результат влияет также длина ресничек, их изгибы, а также цвет. Какие же существуют эффекты при наращивании и как их добиться?
На первый взгляд кажется, что наращённые ресницы не отличаются между собой. Однако применение различных техник материалов и видов ресничек дает абсолютно разнообразные эффекты. На конечный результат влияет также длина ресничек, их изгибы, а также цвет. Какие же существуют эффекты при наращивании и как их добиться?Эффект наращивания ресниц Классика или натуральное
Чтобы нарастить реснички их приклеивают между собственных с помощью специального клея для ресниц. Результат будет зависеть от диаметра и изгиба волосков. Повторимся, наиболее популярным является классическое наращивания, когда на подаренные природой ресницы наклеиваются синтетические. Такой вариант приклеивается по естественному росту волосков. Мастер должен наклеить реснички короткого типа по краям, а в середине чуть длиннее волоски. Натуральный эффект подчеркнёт естественную красоту хозяйки и не утяжеляет общий образ и макияж. Для того, чтобы создать натуральный эффект чаще всего выбираются ресницы шёлкового или норкового типа. Для девушек со светлыми волосами лучше выбрать коричневые волоски, для брюнеток можно использовать более тёмные модели, такие как норка. Натуральный классический эффект также придаст распахнутость взгляду и подчеркнет его красоту. Если добавить для классического эффекта немного больше волосков, то создастся вид ресниц, которые покрашены тушью. Также такой вариант относится к методу наращивание ресниц 2D (2Д).
Для девушек со светлыми волосами лучше выбрать коричневые волоски, для брюнеток можно использовать более тёмные модели, такие как норка. Натуральный классический эффект также придаст распахнутость взгляду и подчеркнет его красоту. Если добавить для классического эффекта немного больше волосков, то создастся вид ресниц, которые покрашены тушью. Также такой вариант относится к методу наращивание ресниц 2D (2Д).Эффект наращивания ресниц Кукольный
Данный эффект создан для тех женщин, которые хотят получить пышный объем ресниц и взгляд, как у куклы. В этом случае волоски искусственного типа имеют одинаковую длину и наносятся по всему росту волос естественных ресниц. Процедура заключается в том, что мастер разделяет волоски на ровные части и в начале идёт наращивание центральных волосков, а затем наносятся реснички на края. Кукольный взгляд с помощью наращенных ресниц также можно использовать и в классическом варианте. С помощью такого эффекта можно добиться объёма 2D. Однако кукольный взгляд подойдет не для всех девушек. Он оптимально будет смотреться на глазах, которые имеют форму миндаля, но не подойдет тем девушкам, которые имеют узкие или выпуклые глаза. Цвет ресниц при кукольном взгляде может быть любой. Однако эффектнее он будет смотреться, если выбрать чёрные искусственные волоски. Чаще всего для создания кукольного эффекта применяются ресницы норка, а их изгиб должен быть больше, чем изгиб собственных естественных волосков.
Однако кукольный взгляд подойдет не для всех девушек. Он оптимально будет смотреться на глазах, которые имеют форму миндаля, но не подойдет тем девушкам, которые имеют узкие или выпуклые глаза. Цвет ресниц при кукольном взгляде может быть любой. Однако эффектнее он будет смотреться, если выбрать чёрные искусственные волоски. Чаще всего для создания кукольного эффекта применяются ресницы норка, а их изгиб должен быть больше, чем изгиб собственных естественных волосков.
Эффект наращивание ресниц 3D или 3Д
Данный эффект схож с первым классическим вариантом, однако для этого мастер применяет утолщенные реснички. Техника нанесения ресниц такая же, как и при классике. Посередине наклеиваются более длинные, а по краям короткие волоски. Для создание накрашенных ресниц мастера используют шёлковые волоски, которые имеют глянцевые блеск и придают дополнительный объем.
Такой эффект подойдет для тех дам, которые постоянно наносят макияж и хотят сэкономить своё время, например, утром перед выходом на работу. Эффект накрашенных ресниц 3д будет держаться долго, если при этом использовались качественные материалы.
Эффект накрашенных ресниц 3д будет держаться долго, если при этом использовались качественные материалы.
Эффект наращивание ресниц Лиса или Лисий
Такой эффект является вторым по популярности после натурального. Принцип основан на том, что пышные и объёмные реснички наклеиваются в уголок глаз. Взгляд при этом получается очень соблазнительным, выразительным и чуть удлиненным.
Лисий эффект делится на следующие варианты:
- Плавный. Это легкий переход от длинны 6-7 B, C на внутреннем угле, в середине длина доходит до 11-12 В, С, D и уменьшается во внешнем угле до 9-8 B, C.
- Резкий переход. В нем, как и в первом варианте, имеется постепенный переход от коротких ресничек 7-8 С, D, L внутреннего угла глаза, но затем резкий переход к самым длинным волоскам и экстремальным изгибам 12-14 D, L во внешних углах.
Для того, чтобы создать лисий взгляд необходимо мысленно или на патче разделить ресницы на четыре части.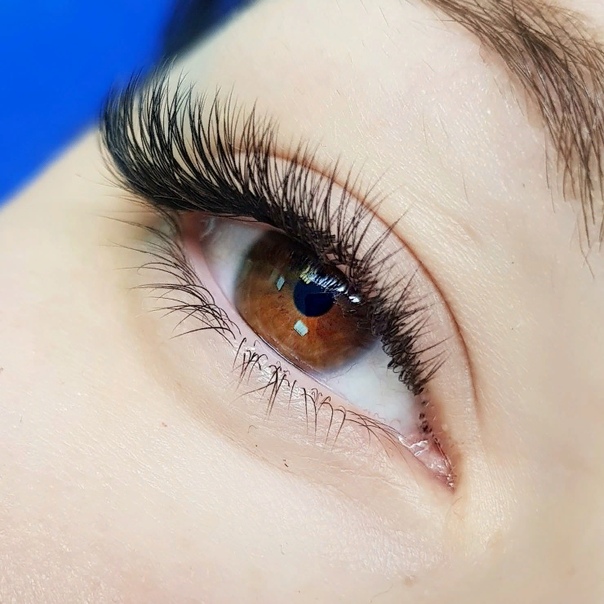 Длинные волоски, которые имеют размер и изгиб 12-14 D, L, закрепить на внешнем угле глаза. Эффект подойдет тем девушкам, которые имеют выпуклую или округлую форму глаз, но не подойдет для тех, к кого глаза далеко посажены. Для лисьего эффекта необходимо выбрать ресницы которые имеют изгиб B, C, D или L.
Длинные волоски, которые имеют размер и изгиб 12-14 D, L, закрепить на внешнем угле глаза. Эффект подойдет тем девушкам, которые имеют выпуклую или округлую форму глаз, но не подойдет для тех, к кого глаза далеко посажены. Для лисьего эффекта необходимо выбрать ресницы которые имеют изгиб B, C, D или L.
Эффект наращивание ресниц Беличий
Данный вариант отличается от остальных тем, что необходимо плавно увеличивать размер волосков. На круглые глаза мастер наносит небольшое количество коротких волосков. Такой эффект поможет открыть взгляд и немножко поднять уголки век. На внутренней стороне века наносятся короткие волоски одинаковой длины, а затем снова короткие. Такой эффект подойдет тем дамам, которые имеют нависшие веки и опущенные уголки глаз, но он неэффективен для тех, у кого глаза далеко расставлены.
Эффект наращивание ресниц Кошачий взгляд
Эффект достаточно сложен по технике и не все лешмейкеры могут его выполнить. Техника кошачьего взгляда состоит в том, что мастер должен вытянуть наружный угол глаза и придать ему округлую форму. Реснички должны плавно увеличиваться с наружным уголком глазом, на край наносятся короткие волоски. Для того, чтобы создать округлую форму века необходимо чередовать изгибы С и В. Если правильно и качественно выполнить наращивание ресниц стрелка, получается очень необычный вариант, который точно создаст яркий образ для любой обладательницы.
Техника кошачьего взгляда состоит в том, что мастер должен вытянуть наружный угол глаза и придать ему округлую форму. Реснички должны плавно увеличиваться с наружным уголком глазом, на край наносятся короткие волоски. Для того, чтобы создать округлую форму века необходимо чередовать изгибы С и В. Если правильно и качественно выполнить наращивание ресниц стрелка, получается очень необычный вариант, который точно создаст яркий образ для любой обладательницы.
Эффект наращивание ресниц Мокрый эффект
Вариант оптимален для тех девушек, которые имеют густые длинные ресницы. В этом случае волоски искусственного типа наносится на определенных расстояниях. В начале идут три волоска естественных и затем наносятся искусственные. С помощью такой комбинации достигается распахнутый взгляд, который точно привлечет к себе внимание.
Эффект наращивание ресниц Лучики
Такой вариант наращивания состоит в том, что искусственные реснички имеют большую длину, чем натуральные. На определенном расстоянии наклеиваются удлинённые волоски. В итоге достигается эффект аккуратного, но естественного взгляда. Ресницам придаётся дополнительный объем. Параметры изгиба и толщины ресничек зависят от индивидуальных особенностей клиента.
На определенном расстоянии наклеиваются удлинённые волоски. В итоге достигается эффект аккуратного, но естественного взгляда. Ресницам придаётся дополнительный объем. Параметры изгиба и толщины ресничек зависят от индивидуальных особенностей клиента.
Цветные ресницы для наращивания
Такой эффект подойдет для особых случаев, а также для экстравагантных девушек, которые любят яркие образы и необычные цветовые сочетания. В этом случае мастер может приклеить абсолютно различные варианты цветных ресниц одного цвета или чередовать цветовую гамму по желанию клиента. Цветные волоски могут быть абсолютно различного цвета, а также иметь смешанные комбинации. В этом случае также можно осуществить наращивание по методу пучков.
Где приобрести ресницы для наращивания?
Не важно, какой эффект для наращивания Вы выбираете: наращивание ресниц Братиславская Катерина или наращивание ресничек классика, все необходимые инструменты и расходные материалы, Вы сможете купить недорого в нашем интернет магазине ВОЛГТЕК. У нас представлен большой выбор товаров для салонов красоты по выгодным ценам и отличного качества.
У нас представлен большой выбор товаров для салонов красоты по выгодным ценам и отличного качества.
Наращивание ресниц — Студия красоты и здоровья «Сирень»
Цена: от 2000 до 2500 р.
Длительность процедуры: 3 — 4 часа.
С помощью наращенных ресниц вы сможете стать обладательницей самых красивых, длинных и пушистых ресниц. Теперь уже Вам не нужно будет тратить время на ежедневные подкрашивания ресниц тушью и прочими средствами, и беспокоиться об их внешнем виде. По крайней мере, на протяжении месяца и даже более Вы сможете наслаждаться своим бесподобным внешним видом. По истечении определенного времени мастер по наращиванию ресниц проведет небольшую их коррекцию, после чего Вы сможете продолжать радовать своими фантастическими ресницами себя и окружающих. Повторное посещение мастера-специалиста займет значительно меньше времени по сравнению с первым посещением. Реснички можно наращивать, готовясь к особому событию или перед отпуском, но большинство наших клиенток носит их постоянно, периодически корректируя. От такой красоты очень трудно отказаться!
От такой красоты очень трудно отказаться!
Процедура наращивания представляет собой процесс прикрепления искусственных ресничек с помощью специального клея, изготовленного из натуральных компонентов, на природные ресницы. Можно удлинить каждую ресничку или выборочно в зависимости от желаемой густоты и эффекта.
При наращивании ресниц определенным образом получаются разные эффекты, вот самые распространенные из них.
Натуральный эффект. Классическое наращивание, создающее ресницы натурального объема. Ресницы одной длины и толщины наращиваются от внешнего угла глаза к внутреннему. Реснички подбираются исходя из состояния натуральных ресниц. Наиболее востребованная длина ресниц при таком эффекте — 8, 10 и 11 мм — они позволяют создать естественный эффект длинных от природы ресниц. На самом деле натуральные ресницы длиной 10 мм пусть нечасто, но встречаются. Сюда же можно включить и обычный эффект, при создании которого используются ресницы длиной 6 и 8 мм, то есть самые распространенные природные длины.
Уголки глаз. Пользуется спросом наращивание ресниц в уголках глаз с внешней стороны. В этом случае создается неповторимый эффект как бы отдельно растущих ресниц, когда контрастируют участки с короткими и длинными ресницами. При этом появляется неотразимый и естественный вид, который только усиливается при наличии одинакового оттенка искусственных и натуральных ресниц.
Лисий эффект. Потрясающий эффект создает наращивание под названием лисий взгляд. В этом случае наращиваются реснички, размер которых увеличиваются к внешнему краю глаза. Этот метод делает глаза немного раскосыми и удлиненными с внешней стороны и придает взгляду особую выразительность и загадочность.
Беличий эффект. Ещё один эффект, позволяющий придать взгляду особую чувственность и загадочность- это так называемый беличий эффект. Сам по себе эффект довольно необычен. Это наращивание с удлинением к внешнему углу глаза, похожее на лисий эффект с той разницей, что при процедуре наращивание ресниц беличий эффект создается путем приклеивания ультрадлинных ресниц не на сам уголок глаза, а с отступом к внутренней стороне примерно 5мм. Очень хороший эффект для вечеринки, хотя, в принципе, подходит и для постоянной носки, особенно ультрамодным женщинам.
Очень хороший эффект для вечеринки, хотя, в принципе, подходит и для постоянной носки, особенно ультрамодным женщинам.
Наращивание премиум класса (3D эффект) достигается благодаря использованию новых современных материалов премиум класса производства США. Это глянцевые блестящие реснички, они немного толще обычных ресниц и темнее по цвету, что делает взгляд более тёмным и выразительным.
Мультицветной эффект — это соединение одного или нескольких оттенков, выбор которых полностью зависит от предпочтений потенциальной обладательницы таких ресниц. Цветные реснички будут отлично дополнять наращенные или натуральные ресницы черного цвета. Для такого наращивания также используются стразы, цветной песок и перья. Особой популярностью эффект мультицветного наращивания пользуется в преддверии какого-либо торжественного мероприятия, хотя и в повседневной жизни такие реснички не станут лишними.
Кукольный эффект. Оригинальный и креативный эффект, позволяющий женщине выгодно отличаться от других — это так называемый кукольный эффект. Этот вид наращивания делает глаза огромными, а женщина становится похожа на куколку. Эффект создается с помощью особо длинных ресничек, накладываемых исключительно поресничным способом. Эффект будет незаменим, если девушка собралась на дискотеку, праздничный карнавал, конкурс красоты.
Этот вид наращивания делает глаза огромными, а женщина становится похожа на куколку. Эффект создается с помощью особо длинных ресничек, накладываемых исключительно поресничным способом. Эффект будет незаменим, если девушка собралась на дискотеку, праздничный карнавал, конкурс красоты.
Разреженный эффект (лучики) — способ наращивания ресниц, последовательно сочетающий две разные длины. При этом создается эффект отдельных длинных ресниц.
Также мастера называют его эффектом «лучики». Глаза в этом случае выглядят наиболее естественно, поскольку для такого наращивания используются реснички разной длины. Накладываются они поочередно на определенном расстоянии друг от друга, и выглядят натурально — ведь живые ресницы растут неравномерно и такой эффект в точности повторяет их натуральный рост. Идеально подходит для женщин, которые недовольны густотой и длиной собственных ресниц. Это способ немного изменить свой внешний вид. Такие ресницы подходят для повседневной носки.
- Рекомендации:
- Не окрашивать перед процедурой
- Не наращивать перед менструацией за 5 дней.
- С наращенными ресницами следует обращаться аккуратно: не спать лицом в подушку, не тереть глаза.
- Повышенная чувствительность глаз.
- Повышенное слезоотделение.
- Воспаление век.
Разные формы глаз. Какое наращивание подойдет Вам?
Разберем подробнее, какие бывают формы глаз и какой эффект наращивания будет смотреться более выигрышно в том или ином случае.
Миндалевидная форма. Если вы та самая обладательница миндалевидной формы глаз, то для вас нет предела творчеству и экспериментам. Вам подойдет абсолютно любой эффект наращивания. Так что можете сами смело выбирать!
Опущенные уголки глаз. Если у вас опущены внешние уголки глаза, то более выигрышно на вас будет смотреться «беличий» эффект. Он помогает визуально поднять уголок глаза и взгляд будет более ярким и открытым.
Он помогает визуально поднять уголок глаза и взгляд будет более ярким и открытым.
Большие и выпуклые глаза. Обладательницам такой формы глаз рекомендуется носить ресницы наиболее приближенные к натуральному эффекту. Это либо классическое наращивание толщиной ресниц не более 0.15 мм, либо двойной объем ультратонкими ресницами. Так же вам подойдет лисий эффект, который визуально поможет скорректировать форму глаза ближе к миндалевидному.
Близко посаженные глаза. Девушкам с такой формой и разрезом глаз подойдет любой эффект обязательно с загущением внешнего уголка (лисий, беличий). Так же, подобную форму глаз, можно корректировать правильно подобранной длиной ресниц.
Глубоко посаженные глаза. Если у вас глубоко посаженные глаза, то корректировать их следует с помощью техники «округления». В таком случае, основной акцент будет приходиться на цетральную часть глаза, визуально делая взгляд более открытым и широким.
Нависшее веко. Девушкам с нависшими веками или Азиатским разрезом глаз, подойдет наращивание ресницами L/L+ изгибами. Это ресницы, которые имеют более крутой изгиб чем все остальные, что позволяет им быть более заметными на глазах с нависшим веком.
Девушкам с нависшими веками или Азиатским разрезом глаз, подойдет наращивание ресницами L/L+ изгибами. Это ресницы, которые имеют более крутой изгиб чем все остальные, что позволяет им быть более заметными на глазах с нависшим веком.
Заключение
Обладая знаниями о том, какая у вас форма глаз и какой разрез, можно быть уверенной в том, что вместе с вашим мастером, вы добьетесь наилучшего результата в корректировки и придания нужных акцентов непосредственно на ваши глаза. Нарастив реснички, так же не забывайте о правилах ухода за ними, чтобы они радовали вас и окружающих как можно дольше. О том, какие средства лучше использовать при снятии макияжа с нарощенными ресницами и многое другое, вы можете прочитать в статье «Наращивание ресниц: правильный уход».
Наращивание ресниц: модные тренды 2020
19 / 12 / 19
Во внешнем виде всех представительниц прекрасного пола огромное значение отводится ресницам. Это неудивительно, ведь женский взгляд способен на многое: привлекать внимание окружающих и вызывать восхищение у мужчин. В том случае, когда необходимо сделать взгляд более выразительным, на помощь приходят нарощенные ресницы.
В том случае, когда необходимо сделать взгляд более выразительным, на помощь приходят нарощенные ресницы.
Нарощенные ресницы: методики и характеристики процедуры
Тонкие от природы ресницы и невыразительный взгляд с легкостью устраняются с помощью наращивания ресниц. Сущность данной процедуры заключается в том, что к натуральным приклеиваются дополнительные реснички. Мастера используют несколько подходов для наращивания ресниц:
- пучковый;
- поресничный.
Поресничное наращивание отличается наибольшей натуральностью. Максимальный объем и естественная красота ресниц достигаются за счет тонкой работы мастера, в процессе которой к каждой ресничке приклеивается дополнительный волосок. Безусловно, для выполнения поресничного наращивания требуется гораздо больше времени. Эффект будет соответствовать всем ожиданиям. В результате профессионально выполненной процедуры исключается необходимость изменения внешнего вида ресниц за счет других манипуляций. Чаще всего для выполнения наращивания по данному методу используются шелковые и норковые волокна.
Для женщин, которые стремятся изменить объем ресниц, актуальным является вопрос, как наращивают ресницы с помощью пучкового метода. Сущность пучкового метода заключается в том, что дополнительные реснички приклеиваются пучками. Пучки содержат по 3-4 дополнительные реснички. Разница пучкового и поресничного метода заключается в следующем:
- возможность быстро нарастить ресницы;
- доступная стоимость пучкового наращивания;
- максимальная длина и густота ресниц;
- возможность подбора оттенков пучков.
Благодаря большому разнообразию оттенков для наращивания каждая клиентка может подобрать дополнительные реснички, которые являются наиболее подходящими к натуральному цвету. Также существует возможность подобрать оптимальную частоту и длину дополнительных ресниц. Для того чтобы достичь надежного приклеивания, применяется специальный гель, который позволяет закрепить пучки волокон на веке. Для наращивания используются исключительно безопасные материалы, которые не вызывают аллергических реакций.
Эффекты наращивания ресниц
При наращивании ресниц можно получить целый ряд эффектов, которые имеют как отличия, так и сходства. Классическими эффектами являются:
- натуральный;
- кукольный;
- лисий;
- беличий;
- разреженный.
Для того чтобы получить максимально естественный вид ресниц, мастер применяет натуральное классическое наращивание. Его особенность заключается в повторении формы изгиба и длины натуральных ресниц. Не менее популярным является беличий эффект, который отличается наличием коррекционных свойств. Данный эффект рекомендуется тем дамам, у которых внешний угол глаз заметно опущен. С помощью применения такого подхода удается внешне изменить разрез глаз, визуально приподняв внешний угол. Лисий эффект заключается в удлинении ресниц к внешнему углу глаза. Он подойдет для девушек, у которых приподнят внешний угол глаз. Максимальная длина ресниц в данном подходе — возле уголков глаз. Популярность набирает модный эффект разреженных ресниц, в котором сочетаются контрастные по длине реснички, изображающие при этом лучики. Кукольный эффект также является декоративным, его особенность — применение длинных ресничек по всей длине глаза.
Кукольный эффект также является декоративным, его особенность — применение длинных ресничек по всей длине глаза.
Для того чтобы подобрать соответствующий эффект и подход к созданию нарощенных ресниц, следует обратиться к профессиональному мастеру, который оценит состояние ресниц, объем и изгиб на момент наращивания. Наращивание ресниц в салоне красоты Bloom — лучшее решение для получения результата, который подчеркнет природную красоту глаз. Лучшие мастера используют безопасные материалы и профессиональные инструменты для создания шикарного эффекта.
Виды наращивания ресниц, техники, технологии, эффекты, секреты
Густые и длинные ресницы от природы – большая редкость. И даже обладательницы таких красивых ресничек со временем сталкиваются с проблемой их выпадения.
Волоски редеют, а новые вырастают тонкими и короткими. В некоторых случаях, приходится прибегать к различным способам их восстановления, один из которых – наращивание – самый быстрый, однако не всегда самый щадящий способ.
Техники и технологии наращивания
На сегодняшний день существует масса техник и видов наращивания ресниц: пучковое, поштучное, поресничное, объемное, 2д и 3д наращивание ресниц. Каждая школа придумывает новые названия своим техникам: японская, голливудское, европейское.
На самом деле специалисты различают две основные технологии наращивания ресниц поштучное и объемное, все остальные по большому счету всего лишь маркетинг. Всё же давайте рассмотрим какие названия техник можно встретить сегодня в данной сфере услуг и что они из себя представляют:
Японская
Она подразумевает наложение одной искусственной ресницы на одну живую. Такую технологию ещё называют поресничным наращиванием. Используются следующие материалы: норка, соболь, шелк.
Длина искусственных ресничек зависит от желаемого эффекта. Чаще самые длинные материалы приклеиваются со стороны внешнего края и идут с убыванием к внутреннему уголку.
Американская
Данная технология мало чем отличается от японской. Различие состоит лишь в материалах. Здесь используются силиконовые или каучуковые ресницы. Их преимущество состоит во влагостойкости и переносимости перепадов температур. С ними можно спокойно посещать баню, бассейны, плавать в море.
Индонезийская
Суть индонезийской технологии так же заключается в поштучном наращивании, однако при этом на каждую ресницу наносится особый клеящий состав с питательным витаминным комплексом. Это питает корни волосков, обеспечивает сохранность и укрепление натуральных ресничек.
Объемная технология
Интересный метод для создания объема от 2D до 5D и даже более. Здесь на каждую ресницу клеится по несколько искусственных. Но главное преимущество в том, что после наращивания эффект натуральности сохраняется за счет использования самых тонких материалов (0,07 мм).
Пучковое наращивание
Эта технология занимает малое количество времени при наращивании, но и эффект длится относительно не долго. Для наращивания используются пучки, состоящие из нескольких ресниц спаянных с одной стороны. Пучки приклеиваются на некотором расстоянии друг от друга.
Для наращивания используются пучки, состоящие из нескольких ресниц спаянных с одной стороны. Пучки приклеиваются на некотором расстоянии друг от друга.
Виды наращивания ресниц
Прежде чем перейти к описанию поэтапного процесса, рассмотрим подробнее, какие существуют на сегодняшний день основные виды наращивания ресниц. Ведь у каждой девушки свой индивидуальный тип лица. И крайне важно подобрать для себя то, что подойдет лучше всего и будет смотреться естественно и красиво.
Наращивание уголков
Данный вид предполагает приклеивание ресниц от внешнего края и только до середины глаза. Если натуральные реснички светлые, тогда им требуется предварительная покраска, поскольку тушью пользоваться после наращивания не рекомендуется.
Неполное наращивание
Такой вид подойдёт обладательницам длинных от природы ресниц. С его помощью добавляется объём. Искусственный материал подбирается по размеру максимально приближённо к натуральным волоскам.
Ресницы приклеиваются на небольшом расстоянии друг от друга. Здесь, как и при наращивании уголков, может потребоваться предварительная покраска во избежание резкого контраста после процедуры.
Полное наращивание
Это и есть полноценное поресничное наращивание — самый изящный, практичный и естественный вид. Если всё сделано профессионально и правильно, то коррекция может не потребоваться на протяжении 2-3 месяцев.
Наращивание в 2 ряда (театральный или 3D эффект)
Этот вид подходит для смелых и ярких людей, желающих сделать акцент на глазах. Это такой же поресничный метод, только на каждую родную ресничку приклеивается по 2 искуственных.
В итоге получается очень эффектный взгляд и хороший объём. Также этот вид мастера советуют девушкам с малым количеством своих ресниц.
Виды волосков
Виды варьируются в зависимости от толщины и плотности:
- Норка. Эти волоски самые лёгкие и тонкие.
 Чаще их используют, если родные ресницы в плохом состоянии (ломкие, поврежденные). Иначе другие искусственные материалы только усугубят ситуацию.
Чаще их используют, если родные ресницы в плохом состоянии (ломкие, поврежденные). Иначе другие искусственные материалы только усугубят ситуацию. - Шёлк. Они немного толще и являются довольно пушистыми. Такие волоски идеально подходят для создания естественного объемного эффекта.
- Соболь – самые длинные, пушистые, и, соответственно, достаточно «тяжелые».
 Чаще их используют, если родные ресницы в плохом состоянии (ломкие, поврежденные). Иначе другие искусственные материалы только усугубят ситуацию.
Чаще их используют, если родные ресницы в плохом состоянии (ломкие, поврежденные). Иначе другие искусственные материалы только усугубят ситуацию.Кстати, название волосков не имеет ничего общего с натуральными материалами (шёлком или мехом соболя). Они производятся из синтетических волокон. А такое название получили благодаря своим характеристикам.
Так же они производятся различной формы. В зависимости от изгиба классифицируются и обозначаются так:
- B – прямые волоски;
- C – изогнутые;
- CC/B – сильный изгиб;
- L – изгиб приходится на край волоска.
Эффекты, достигаемые наращиванием
Все эффекты подбираются, как правило, индивидуально для каждой клиентки с учетом возраста, формы, цвета глаз и статуса. Некоторые из них могут применяться только для определенных торжественных случаев и снимаются на следующий день после их окончания, так как неуместны для повседневного ношения.
Некоторые из них могут применяться только для определенных торжественных случаев и снимаются на следующий день после их окончания, так как неуместны для повседневного ношения.
Обычный эффект
Для его создания используются самые короткие волоски (6 или 8 мм). Они берутся одной длины. Внешне трудно отличить от своих ресниц, но прибавляется небольшой объём.
Натуральный
Данный эффект является наиболее популярным и универсальным. Здесь используются материалы двух длин (6 и 8 мм или 8 и 10 мм). Реснички разных длин клеятся поочередно. В итоге взгляд приобретает выразительность и шарм.
Лисий эффект — лисий взгляд
Для лисьего взгляда потребуются материалы трёх длин. Сначала во внешнем уголке чередуются реснички двух самых больших размеров и постепенно в ход идут короткие волоски.
Кукольный
Это очень интересный эффект, для которого используются самые длинные волоски (12-15 мм). Одинаковые реснички клеятся даже у внутреннего уголка. Выглядит очень необычно и притягательно, но не совсем подходит для ежедневной носки.
Выглядит очень необычно и притягательно, но не совсем подходит для ежедневной носки.
Беличий
Здесь можно добиться визуального поднятия внешнего края глаза. Реснички подбираются двух длин – очень длинные и очень короткие. Сначала клеятся длинные волоски на внешней части глаза.
При этом нужно отступить полсантиметра от края, так обрабатывается около 1 см линии роста ресниц, затем клеятся короткие волоски на оставшуюся часть.
Коктейльный
Это эффект чередования. Есть две вариации: чередование по длине либо по толщине. Например, поочередно по одной штуке приклеивается сначала длинная, затем короткая волосинка. Либо сначала «соболь», затем «норка».
Разреженный эффект
Это довольно интересный способ. Выбирается материал совсем разной длины (возьмём для примера 8 и 12 мм). Реснички 12 мм приклеиваются через определённое количество коротких волосков. То есть можно клеить длинные ресницы через каждые 5 коротких. Длинные волоски можно сделать цветными.
Мультицветной
Здесь всё полностью зависит от фантазии мастера и клиента. Можно использовать все возможные цветовые гаммы. Например, взять за основу натуральный эффект, но брать ресницы разных цветов (сделать плавный переход от красного к синему цвету).
Как проходит процедура
Классическая процедура наращивания состоит как правило из следующих этапов:
- Очищенные от косметики ресницы протираются специальным обезжиривающим средством;
- Подбирается длина и материал волосков, технология и желаемый эффект от наращивая;
- Разделяются верхние и нижние ресницы с помощью специально предназначенных наклеек. Это необходимо во избежание склеивания в процессе процедуры;
- С помощью пинцета основание каждой синтетической реснички обмакивается в клей или смолу для наращивания и помещается на родной волосок.
Если клеевого состава на материале слишком много, то следует удалить его излишки. Важно располагать искусственный материал ровно – ресничка на ресничку. И так до получения конечного результата.
И так до получения конечного результата.
Сколько времени занимает процесс
У опытного мастера в среднем на наращивание уходит от 1,5 до 2 часов (у новичков чуть больше). Если производить наращивание самой себе, или в домашних условиях, соответственно на это уйдёт гораздо больше времени.
Во избежание травм и нежелательных последствий мы рекомендуем выполнять данную процедуру в специализированных салонах у квалифицированных специалистов.
Редокс-регуляция подвижных ресничек при заболеваниях дыхательных путей
Реферат
Подвижные реснички на клетках дыхательных путей необходимы для выведения захваченных слизью частиц из легких. Ресничные эпителиальные клетки дыхательных путей подвергаются уникальному воздействию окислителей за счет улавливания частиц, мусора и патогенов в слизи и прямого воздействия вдыхаемых газов-окислителей. Динеин-АТФазы, двигатели, управляющие подвижностью ресничек, чувствительны к локальной окислительно-восстановительной среде внутри каждой реснички. Некоторые редокс-чувствительные белки, локализованные в ресничках, модулируют активность динеина и включают протеинкиназу А, протеинкиназу С и протеинфосфатазу 1.Более того, реснички богаты известными белками, регулирующими окислительно-восстановительный потенциал, и белками, содержащими тиоредоксиновый домен, которые имеют решающее значение для поддержания сбалансированной окислительно-восстановительной среды. Важно отметить, что нонсенс-мутация в TXNDC3, которая содержит мотив тиоредоксина, была недавно идентифицирована как вызывающая заболевание при первичной цилиарной дискинезии, наследственном заболевании подвижных ресничек, приводящем к нарушению мукоцилиарного клиренса. Здесь мы рассматриваем современное понимание роли (-ов) окислителей в изменении функции ресничек дыхательных путей.Мы фокусируемся на окислителях, образующихся в дыхательных путях, окислительно-восстановительных мишенях ресничек, которые модулируют биение ресничек, и дисбалансе окислительно-восстановительного состояния, влияющем на здоровье и болезни.
Некоторые редокс-чувствительные белки, локализованные в ресничках, модулируют активность динеина и включают протеинкиназу А, протеинкиназу С и протеинфосфатазу 1.Более того, реснички богаты известными белками, регулирующими окислительно-восстановительный потенциал, и белками, содержащими тиоредоксиновый домен, которые имеют решающее значение для поддержания сбалансированной окислительно-восстановительной среды. Важно отметить, что нонсенс-мутация в TXNDC3, которая содержит мотив тиоредоксина, была недавно идентифицирована как вызывающая заболевание при первичной цилиарной дискинезии, наследственном заболевании подвижных ресничек, приводящем к нарушению мукоцилиарного клиренса. Здесь мы рассматриваем современное понимание роли (-ов) окислителей в изменении функции ресничек дыхательных путей.Мы фокусируемся на окислителях, образующихся в дыхательных путях, окислительно-восстановительных мишенях ресничек, которые модулируют биение ресничек, и дисбалансе окислительно-восстановительного состояния, влияющем на здоровье и болезни. Наконец, мы рассматриваем модели заболеваний, таких как курение, астма, употребление алкоголя и инфекции, а также прямое применение оксидантов, которые влияют на окислительно-восстановительный баланс как модулятор подвижности ресничек.
Наконец, мы рассматриваем модели заболеваний, таких как курение, астма, употребление алкоголя и инфекции, а также прямое применение оксидантов, которые влияют на окислительно-восстановительный баланс как модулятор подвижности ресничек.
Ключевые слова
S -нитрозирование
Спирт
Реснички
Регуляция окислительно-восстановительного потенциала
Пероксид водорода
Супероксид
Оксид азота
Сокращения 9000TPOK5
, вызываемый алкоголем,
, вызываемый алкоголем частота биений ресничек
DTNB5,5′-дитио-бис- [2-нитробензойная кислота]
FiO 2фракция вдыхаемого кислорода
Hsp90белок теплового шока 90
l-NAMEМетиловый эфир N-нитроаргинина
NOXникатиномид-аденин-динуклеотид-фосфат-оксидаза
Nrf2Ядерный фактор (эритроидный 2) -подобный 2
PCDпервичная цилиарная дискинезия
PP2Aпротеинфосфатаза 2A
RHAMM рецептор RHAMM, связанный с рецептором азотадля гиалуроновой кислоты
для гиалуроновой кислоты
виды
RSVреспираторно-синцитиальный вирус
sGCрастворимая гуанилилциклаза
TrxR2 900 04 тиоредоксинредуктаза 2 TXNDC, содержащий тиоредоксиновый домен
XDH / XOксантиндегидрогеназа / ксантиноксидаза
Рекомендуемые статьиЦитирующие статьи (0)
Просмотреть аннотацию © 2019 Авторы. Опубликовано Elsevier B.V.
Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирование статей
Структурная основа того, как движутся реснички ритма | Бионаука
Аннотация
Подвижная ресничка — это чудо механики, клеточная наномашина, которая производит высокоскоростные биения на основе цикла изгибов, которые движутся вдоль аксонемы, состоящей из 9 + 2 микротрубочек. Молекулярные двигатели, динеины, приводят в действие биение ресничек. Динеины компактируются во внутреннюю и внешнюю ветви динеина, активность которых строго регулируется, вызывая скольжение микротрубочек и изгиб аксонемы.Гипотеза точки переключения была разработана давно, чтобы объяснить, как скольжение в присутствии взаимодействий аксонемной радиальной спицы и центральной пары вызывает биение ресничек. С тех пор новая генетическая, биохимическая и структурная сложность была обнаружена, в частности, с мутантами Chlamydomonas, с помощью высокоскоростного анализа движения с высоким разрешением и криоэлектронной томографии. Мы стоим на пороге новых открытий, касающихся молекулярного контроля подвижности, которые расширяют и уточняют наше понимание основных событий, лежащих в основе переключения активности рук, а также формирования и распространения изгибов.
Мы стоим на пороге новых открытий, касающихся молекулярного контроля подвижности, которые расширяют и уточняют наше понимание основных событий, лежащих в основе переключения активности рук, а также формирования и распространения изгибов.
Подвижные реснички — самая ранняя из известных клеточных органелл , обнаруженная около 1647 года, когда ван Левенгук впервые увидел ресничные протисты в свой микроскоп. Хотя вопрос о механизме движения ресничек с того времени интересовал биологов и был предметом значительных спекуляций (Gray 1928), только с зарождением биологической электронной микроскопии в середине двадцатого века и появлением современной клеточной биологии несколько спустя годы появилась правдоподобная модель.Ключом к модели было понимание ультраструктуры ресничек. Исследования нескольких исследователей (Manton 1953, Fawcett and Porter 1959, Afzelius 1959, Gibbons IR и Grimstone 1960, Gibbons IR 1961) установили универсальность подвижной аксонемы ресничек 9 + 2 и ее важных субструктурных элементов, включая радиальные спицы и руки. , которые, как показали Гиббонс И.Р. и Роу (1965), были цилиарными АТФазами, которые они назвали динеинами . Типичное изгибное движение ресничек и волнообразное движение биения жгутиков показано на рисунке 1.Рисунок был адаптирован на основе результатов количественного анализа прямого и обратного плавания (Brokaw and Kamiya, 1987). Chlamydomonas . На рисунке 2 показана ранняя электронная микрофотография образца 9 + 2 жаберных ресничек мидии, стандартного экспериментального организма для ранних исследований ресничек (Warner and Satir 1974), а также диаграмма, показывающая ось структуры 9 + 2 относительно плоскость изгиба.
, которые, как показали Гиббонс И.Р. и Роу (1965), были цилиарными АТФазами, которые они назвали динеинами . Типичное изгибное движение ресничек и волнообразное движение биения жгутиков показано на рисунке 1.Рисунок был адаптирован на основе результатов количественного анализа прямого и обратного плавания (Brokaw and Kamiya, 1987). Chlamydomonas . На рисунке 2 показана ранняя электронная микрофотография образца 9 + 2 жаберных ресничек мидии, стандартного экспериментального организма для ранних исследований ресничек (Warner and Satir 1974), а также диаграмма, показывающая ось структуры 9 + 2 относительно плоскость изгиба.
Рисунок 1.
Паттерны изгиба цилиарного (а) и жгутикового (б) движения у Chlamydomonas, зафиксированные с помощью высокоскоростной фотосъемки со вспышкой.Рисунки иллюстрируют изгибные паттерны движения, типичные для эпителиальных ресничек, и волнообразные паттерны движения, типичные для изгиба жгутиков эукариот. Серые стрелки указывают направление движения клеток. Черная полоса у основания ресничек или жгутиков имеет длину 5 микрометров и отмечает место прикрепления ресничек или жгутиков к клетке. Открытая стрелка на панели (а) указывает направление переднего изгиба ресничек. Открытая стрелка на панели (b) указывает направление распространения изгиба жгутика.Адаптировано с разрешения Brokaw и Kamiya (1987).
Черная полоса у основания ресничек или жгутиков имеет длину 5 микрометров и отмечает место прикрепления ресничек или жгутиков к клетке. Открытая стрелка на панели (а) указывает направление переднего изгиба ресничек. Открытая стрелка на панели (b) указывает направление распространения изгиба жгутика.Адаптировано с разрешения Brokaw и Kamiya (1987).
Рисунок 1.
Паттерны изгиба цилиарного (a) и жгутикового (b) движения у Chlamydomonas, зафиксированные с помощью высокоскоростной фотосъемки со вспышкой. Рисунки иллюстрируют изгибные паттерны движения, типичные для эпителиальных ресничек, и волнообразные паттерны движения, типичные для изгиба жгутиков эукариот. Серые стрелки указывают направление движения клеток. Черная полоса у основания ресничек или жгутиков имеет длину 5 микрометров и отмечает место прикрепления ресничек или жгутиков к клетке.Открытая стрелка на панели (а) указывает направление переднего изгиба ресничек. Открытая стрелка на панели (b) указывает направление распространения изгиба жгутика. Адаптировано с разрешения Brokaw и Kamiya (1987).
Адаптировано с разрешения Brokaw и Kamiya (1987).
Рис. 2.
Поперечный разрез подвижной реснички. (а) Электронная микрофотография единственной реснички из латеральных ресничек жабр моллюска при взгляде изнутри клетки по направлению к дистальному концу реснички. Дублет 1 расположен в положении на 12 часов, а другие внешние микротрубочки дублета пронумерованы по часовой стрелке в направлении, в котором указывают плечи динеина.Также проиллюстрирован мост 5–6. Латеральные реснички бьются в плоскости, определяемой дублетом 1 и мостиком 5-6, и в этих ресничках эффективное или прямое направление изгиба направлено к мосту 5-6. Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, иллюстрирующая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее динеиновые рукава, проекции центральной пары, мостик 5-6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Рис. 2.
Поперечный разрез подвижной реснички. (а) Электронная микрофотография единственной реснички из латеральных ресничек жабр моллюска при взгляде изнутри клетки по направлению к дистальному концу реснички. Дублет 1 расположен в положении на 12 часов, а другие внешние микротрубочки дублета пронумерованы по часовой стрелке в направлении, в котором указывают плечи динеина. Также проиллюстрирован мост 5–6. Латеральные реснички бьются в плоскости, определяемой дублетом 1 и мостиком 5-6, и в этих ресничках эффективное или прямое направление изгиба направлено к мосту 5-6.Источник: перепечатано с разрешения Warner and Satir (1974). (b) Диаграмма, иллюстрирующая основные особенности аксонемы 9 + 2 из подвижных ресничек многоклеточных животных, включая микротрубочки наружного дублета и центральной пары, внешнее и внутреннее динеиновые рукава, проекции центральной пары, мостик 5-6, радиальные спицы, и регуляторный комплекс динеина и связи нексина.
Форма ресничного биения хорошо изучена. Хотя иногда он может сильно варьироваться, даже в одной и той же клетке, типичный толчок ресничек обычно представляет собой короткий, очевидно жесткий эффективный ход, при котором ресничка указывает в направлении генерирования силы, и гибкий ход восстановления, при котором кончик медленнее отводится назад (рисунки 1а и 3), тогда как типичные сперматозоиды с 9 + 2 хвостами ресничек (называемые жгутиками ) бьют волнообразными движениями (рисунок 1b).И изгибное, и волнообразное движение (рисунок 1) вызывают поток жидкости, перемещение воды или слизи по реснитчатой поверхности (рисунок 1a) или продвижение реснитчатых клеток (рисунок 1b) через воду со скоростью примерно до 1 миллиметра в секунду, в зависимости от таких параметров, как длина ресничек и частота биений. Почти все реснички работают в гидродинамическом режиме с низким числом Рейнольдса, в котором силы вязкости имеют первостепенное значение, а силами инерции можно пренебречь (Holwill 1974). По сути, эффективный ход при изгибном движении (открытая стрелка на рисунке 1а) перемещается перпендикулярно поверхности ячейки, что вызывает скорость жидкости, примерно вдвое превышающую скорость хода восстановления, которая движется более параллельно поверхности.
Хотя иногда он может сильно варьироваться, даже в одной и той же клетке, типичный толчок ресничек обычно представляет собой короткий, очевидно жесткий эффективный ход, при котором ресничка указывает в направлении генерирования силы, и гибкий ход восстановления, при котором кончик медленнее отводится назад (рисунки 1а и 3), тогда как типичные сперматозоиды с 9 + 2 хвостами ресничек (называемые жгутиками ) бьют волнообразными движениями (рисунок 1b).И изгибное, и волнообразное движение (рисунок 1) вызывают поток жидкости, перемещение воды или слизи по реснитчатой поверхности (рисунок 1a) или продвижение реснитчатых клеток (рисунок 1b) через воду со скоростью примерно до 1 миллиметра в секунду, в зависимости от таких параметров, как длина ресничек и частота биений. Почти все реснички работают в гидродинамическом режиме с низким числом Рейнольдса, в котором силы вязкости имеют первостепенное значение, а силами инерции можно пренебречь (Holwill 1974). По сути, эффективный ход при изгибном движении (открытая стрелка на рисунке 1а) перемещается перпендикулярно поверхности ячейки, что вызывает скорость жидкости, примерно вдвое превышающую скорость хода восстановления, которая движется более параллельно поверхности. Гидромеханические аспекты движения ресничек были тщательно проанализированы и смоделированы (напр., Blake and Sleigh 1974, Smith DJ et al. 2007).
Гидромеханические аспекты движения ресничек были тщательно проанализированы и смоделированы (напр., Blake and Sleigh 1974, Smith DJ et al. 2007).
Рисунок 3.
Диаграммы, иллюстрирующие (а) фазы изгиба ресничек и (б) геометрию скользящей модели микротрубочек для изгиба ресничек. Детали каждой диаграммы обсуждаются в тексте и иллюстрируют геометрию скольжения микротрубочек для эффективных и обратных изгибов микротрубочек, закрепленных в базальном теле и свободно скользящих на дистальной аксонеме.Модель основана на открытии того факта, что микротрубочки нерастяжимы и что их изгибы имеют форму дуг окружности. Источник: перепечатано с разрешения Satir (1968).
Рисунок 3.
Диаграммы, иллюстрирующие (а) фазы изгиба ресничек и (б) геометрию скользящей модели микротрубочек для изгиба ресничек. Детали каждой диаграммы обсуждаются в тексте и иллюстрируют геометрию скольжения микротрубочек для эффективных и обратных изгибов микротрубочек, закрепленных в базальном теле и свободно скользящих на дистальной аксонеме. Модель основана на открытии того факта, что микротрубочки нерастяжимы и что их изгибы имеют форму дуг окружности. Источник: перепечатано с разрешения Satir (1968).
Модель основана на открытии того факта, что микротрубочки нерастяжимы и что их изгибы имеют форму дуг окружности. Источник: перепечатано с разрешения Satir (1968).
В контексте 9 + 2 аксонемных механизмов не существует выдающихся различий между органеллами, первоначально названными ресничками и теми, которые называются жгутиками . Ранние модели предполагали, что для возникновения инсульта девять дублетных микротрубочек модели 9 + 2 последовательно сокращаются, и это казалось биофизически правдоподобным, но с растущим пониманием того, что сокращение мышц зависит от скольжения актиновых и миозиновых нитей, люди начали считать, что сокращение ресничек также может быть вызвано скольжением.Какие элементы будут скользить? Когда Афзелиус (1959) впервые увидел руки, он предположил, что они могут вызывать скольжение дублетных микротрубочек.
Satir (1963) подошел к проблеме, попытавшись использовать электронный микроскоп для фиксации структурных изменений внутри реснички по мере изменения формы биений. Биение можно было остановить, капнув раствор четырехокиси осмия на препарат, бьющийся метахронными волнами. Несколько исследователей, начиная с Gelei (1925, цитируется в Satir 1963), зафиксировали метахрональную волну на простейших, улавливая реснички в последовательных положениях биения вдоль длины волны.Сатир (1963, 1965) зафиксировал метахрональную волну латеральных ресничек мидии, впервые зафиксировав с помощью электронной микроскопии направленные в разные стороны реснички во время мазка. До фиксации частота биений ресничек составляла приблизительно 17 герц (Гц), поэтому биение занимало около 60 миллисекунд (мс). Тридцать одну индивидуальную ресничку измеряли на длине волны 11 микрометров (мкм) в фиксированном препарате, так что реснички были захвачены с разностью фаз около 2 мс между соседними ресничками (Satir 1967).
Биение можно было остановить, капнув раствор четырехокиси осмия на препарат, бьющийся метахронными волнами. Несколько исследователей, начиная с Gelei (1925, цитируется в Satir 1963), зафиксировали метахрональную волну на простейших, улавливая реснички в последовательных положениях биения вдоль длины волны.Сатир (1963, 1965) зафиксировал метахрональную волну латеральных ресничек мидии, впервые зафиксировав с помощью электронной микроскопии направленные в разные стороны реснички во время мазка. До фиксации частота биений ресничек составляла приблизительно 17 герц (Гц), поэтому биение занимало около 60 миллисекунд (мс). Тридцать одну индивидуальную ресничку измеряли на длине волны 11 микрометров (мкм) в фиксированном препарате, так что реснички были захвачены с разностью фаз около 2 мс между соседними ресничками (Satir 1967).
Сатир (1963, 1965) предположил, что после такой быстрой фиксации исследование тонкой структуры аксонемы ресничек, захваченных в различных положениях, покажет, как происходит сокращение. Что исследовать, было неочевидно, но затем Roth и Shigenaka (1964) показали, что в некоторых аксонемах кончики не сохраняют паттерн 9 + 2; некоторые из дуплетов стали синглетами. Можно представить себе, что если некоторые из дублетов сократятся, чтобы согнуть ресничку, скажем, в направлении эффективного удара, сокращенные дублеты станут синглетами и исчезнут с кончиков раньше, чем дублеты на противоположной стороне (на стороне восстановления). Инсульт).Затем, когда ресничка изгибалась в направлении восстановительного удара, был бы виден прямо противоположный результат: дублеты, которые сокращались и исчезали, были бы теми, которые были на стороне восстановительного удара, тогда как те, которые находились на стороне эффективного удара, сохранялись.
Что исследовать, было неочевидно, но затем Roth и Shigenaka (1964) показали, что в некоторых аксонемах кончики не сохраняют паттерн 9 + 2; некоторые из дуплетов стали синглетами. Можно представить себе, что если некоторые из дублетов сократятся, чтобы согнуть ресничку, скажем, в направлении эффективного удара, сокращенные дублеты станут синглетами и исчезнут с кончиков раньше, чем дублеты на противоположной стороне (на стороне восстановления). Инсульт).Затем, когда ресничка изгибалась в направлении восстановительного удара, был бы виден прямо противоположный результат: дублеты, которые сокращались и исчезали, были бы теми, которые были на стороне восстановительного удара, тогда как те, которые находились на стороне эффективного удара, сохранялись.
Чтобы проверить это, необходимо однозначно идентифицировать аксонемные дублеты в поперечном срезе кончика. К счастью, боковые реснички жабр имели особую перемычку между двумя дублетами (рис. 2), которые, казалось, находились в одном и том же положении в каждом поперечном сечении и, по-видимому, идентифицировали дублеты в направлении эффективного удара. Предполагалось, что мостик в этих ресничках был стабильным и что идентифицированные дублеты были дублетами 5 и 6, согласно стандартной нумерации (фиг. 2). Еще одно предположение заключалось в том, что все дублеты имели одинаковую морфологическую длину в прямой ресничке до того, как произошло изгибание.
Предполагалось, что мостик в этих ресничках был стабильным и что идентифицированные дублеты были дублетами 5 и 6, согласно стандартной нумерации (фиг. 2). Еще одно предположение заключалось в том, что все дублеты имели одинаковую морфологическую длину в прямой ресничке до того, как произошло изгибание.
Первоначальная демонстрация (Satir 1965) показала, что дублеты на концах ресничек эффективного наведения и на концах ресничек наведения на восстановление исчезают в разных последовательностях, причем дублеты 4–6 сохраняются в первых после дублетов 9, 1 и 2 ( рядом с 9 в конце последовательности), а дублеты 9, 1 и 2 сохраняются в последнем после того, как 4–6 закончились (рис. 3).Этот результат не был совместим с простой сократительной моделью, как было заявлено, но фактически был полной противоположностью того, что ожидалось. Однако это можно объяснить, если для компенсации изгиба дуплеты скользят друг мимо друга, не меняя длины. Хотя это было спорным в то время, это стало оригинальным доказательством модели подвижности ресничек скользящих микротрубочек.
\ begin {уравнение *} \ Delta l_n = d_n \, \ Sigma \ alpha, \ end {уравнение *}
, где d n — эффективный диаметр аксонемы для дублета n , а Σα — сумма изгибов реснички, измеренная в радианах.Для дублетов 1 и 5, которые находятся на противоположных сторонах аксонемы, d n — это просто диаметр аксонемы, около 0,2 мкм, а ожидаемое значение Δ l на кончике составляет 3,5 нанометра (нм) на градус (°) изгиба. Изгиб, который образуется в начале эффективного хода, составляет около 100 °, что дает максимальное прогнозируемое смещение около 100 нм между соседними дублетами.
Изгиб, который образуется в начале эффективного хода, составляет около 100 °, что дает максимальное прогнозируемое смещение около 100 нм между соседними дублетами.Уравнение может быть проверено последовательным разрезанием кончиков ресничек изогнутых ресничек в фиксированной метахрональной волне и непосредственным измерением Δ l .Каждая секция имела толщину примерно 100 нм. В пределах погрешности измерения, как для эффективных, так и для указывающих на восстановление ресничек, измеренное смещение кончика наконечника соответствовало предсказанному значению (рисунок 3; Satir 1968). Более того, несколько различных паттернов были замечены на концах, которые соответствуют снимкам ресничек с разными полными изгибами вдоль аксонемы во время фиксированного удара. Паттерны кончиков и степень смещения дублетов были позже подтверждены разворачиванием изолированных аксонем ресничек Tetrahymena (Sale and Satir 1976).
Основным возражением против этого анализа является то, что фиксация может вызывать релаксацию сокращенных микротрубочек обратно до их исходной длины. Можно было бы возразить, что это маловероятно, потому что фиксация происходит очень быстро и потому что множественные изменения вдоль аксонемы — которые мы обсудим сейчас — несовместимы с расслаблением. Этот вопрос был однозначно решен Саммерсом и Гиббонсом (1971), которые использовали микроскопию темного поля, чтобы напрямую продемонстрировать скольжение между внешними дублетными микротрубочками.
Можно было бы возразить, что это маловероятно, потому что фиксация происходит очень быстро и потому что множественные изменения вдоль аксонемы — которые мы обсудим сейчас — несовместимы с расслаблением. Этот вопрос был однозначно решен Саммерсом и Гиббонсом (1971), которые использовали микроскопию темного поля, чтобы напрямую продемонстрировать скольжение между внешними дублетными микротрубочками.
К началу 1970-х годов стало ясно, что если ресничную мембрану осторожно удалить с помощью мягкого моющего средства (такого как Triton X-100), образовавшаяся голая аксонема может быть реактивирована, чтобы победить добавлением аденозинтрифосфата (АТФ). . Гиббонс Б. Х. и Гиббонс (1972) показали, что для сперматозоидов морского ежа реактивированный ритм был идентичен биению живой спермы, что означало, что практически весь механизм образования и контроля изгиба находится в аксонеме, что подтверждено многими более поздними исследованиями. эксперименты.Мембрана поддерживала концентрацию АТФ и различных ионов на уровне, необходимом для подвижности. Он также обеспечивает такие сигналы, как цАМФ (циклический аденозинмонофосфат) или ионы кальция (Ca 2+ ), которые воздействуют на белки аксонемы, влияя на поведение ресничек.
Он также обеспечивает такие сигналы, как цАМФ (циклический аденозинмонофосфат) или ионы кальция (Ca 2+ ), которые воздействуют на белки аксонемы, влияя на поведение ресничек.
Саммерс и Гиббонс (1971) использовали этот препарат аксонем, но перед добавлением АТФ их кратковременно переваривали трипсином. Анализ просвечивающей электронной микроскопии показал, что целостность радиальной спицы была нарушена трипсином.Вместо того чтобы снова активировать биение, эти аксонемы теперь раздвигались, в конечном итоге увеличиваясь в восемь или девять раз по длине от исходной аксонемы. Этот результат подтвердил вывод о том, что основным взаимодействием в подвижности ресничек было скольжение дублета, приводимое в действие активностью динеиновой руки, так что руки на любом аксонемном дублете ( n ) производили скольжение соседнего дублета ( n + 1). Очевидно, динеиновые рукава могли активироваться по длине каждого дублета, и почти каждый дублет в аксонеме был способен скользить. Кроме того, считалось, что разница между скольжением и изгибом связана с целостностью радиальных спиц и междублетных звеньев.
Кроме того, считалось, что разница между скольжением и изгибом связана с целостностью радиальных спиц и междублетных звеньев.
Итак, внутри аксонемы есть две системы, необходимые для подвижности ресничек: (1) нечувствительная к трипсину система генерирования силы, основанная на взаимодействиях динеин-микротрубочка, способная генерировать неограниченное изотропное скольжение иногда более 10 мкм на дуплет без систематического изгиба. формирование или распространение и (2) чувствительная к трипсину система управления, в зависимости от спица-центральная оболочка и других взаимодействий, где скользящий максимум около 0.1 мкм на дублет (т.е. между соседними дублетами) строго связан с систематическим образованием и распространением изгиба на высокой частоте (до 100 Гц). Brokaw (1989, 1991) предоставил прямое подтверждение и критический количественный анализ скольжения дублета во время образования изгиба и распространения в волнообразном движении. Относительное движение 40-нм золотых шариков, связанных с обнаженными внешними дублетными аксонемами жгутиков сперматозоидов во время АТФ-реактивированного плавания, согласуется со скользящей моделью взаимодействий дублетов. Вернон и Вулли (2002) смогли увидеть скольжение на кончике в соответствии с моделью точки переключения в сперме млекопитающих.
Вернон и Вулли (2002) смогли увидеть скольжение на кончике в соответствии с моделью точки переключения в сперме млекопитающих.
Форма волны и направление прогрессии изгиба являются эпифеноменами, зависящими от времени скольжения, переключения и управляющих событий в аксонеме и могут изменяться. Главной демонстрацией этого вывода является переход от плавательного изгибающего движения вперед к волнообразному движению, ведущему к обратному плаванию и обратно у жгутиков Chlamydomonas , демонстрирующих реакцию избегания (сравните цилиарный, рисунок 1a, с движением жгутика, рисунок 1b).Это изменение может быть продемонстрировано на моделях клеток Chlamydomonas с демембранными, реактивированными АТФ или жгутиковых аппаратах путем добавления или удаления Ca 2+ из раствора для реактивации (Hyams and Borisy 1978, Kamiya and Witman 1984).
Сейл и Сатир (1977) расширили эксперименты Саммерса и Гиббонса (1971) с помощью анализа просвечивающей электронной микроскопии скользящих аксонем Tetrahymena . После скольжения можно было изучить перекрытие между соседними дублетами.Они обнаружили, что дублет n + 1 всегда располагался наверху дублета n в таких перекрытиях. Они пришли к выводу, что динеиновые рукава имеют однородную полярность, всегда проходя по дублету, к которому они были постоянно прикреплены, к основанию ресничек. С появлением систем для изучения полярности микротрубочек, основание ресничек было определено как отрицательный конец дублета, а аксонемные динеины в совокупности, следовательно, были двигателем с отрицательным концом. Тесты изолированных внешних динеиновых плеч и многих компонентов внутренних динеиновых плеч в анализах in vitro подтвердили этот вывод (Vale and Toyoshima, 1988).
После скольжения можно было изучить перекрытие между соседними дублетами.Они обнаружили, что дублет n + 1 всегда располагался наверху дублета n в таких перекрытиях. Они пришли к выводу, что динеиновые рукава имеют однородную полярность, всегда проходя по дублету, к которому они были постоянно прикреплены, к основанию ресничек. С появлением систем для изучения полярности микротрубочек, основание ресничек было определено как отрицательный конец дублета, а аксонемные динеины в совокупности, следовательно, были двигателем с отрицательным концом. Тесты изолированных внешних динеиновых плеч и многих компонентов внутренних динеиновых плеч в анализах in vitro подтвердили этот вывод (Vale and Toyoshima, 1988).
Следствие однородной полярности активного скольжения означает, что во время биения ресничек только некоторые дублеты или части дублетов имеют активные руки в любой момент времени. Когда дублет n + 1 движется к основанию дублета n , движение является пассивным, а руки дублета n в целом неактивны. Тщательное исследование ресничек жаберных мидий показало, что во время эффективного удара дублеты примерно на половине аксонемы вызывали активное скольжение, тогда как скольжение противоположной половины было пассивным, а во время восстановительного удара половинки менялись местами (Satir and Matsuoka 1989).Во время биения ресничек скользящая активность переключается между двумя половинами аксонемы, определяемыми линией, которая проходит между центральной парой и разделяет мост и дуплет 1 пополам (рис. 2). Эта гипотеза представляет собой модель точки переключения ресничек (см. Также Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
Тщательное исследование ресничек жаберных мидий показало, что во время эффективного удара дублеты примерно на половине аксонемы вызывали активное скольжение, тогда как скольжение противоположной половины было пассивным, а во время восстановительного удара половинки менялись местами (Satir and Matsuoka 1989).Во время биения ресничек скользящая активность переключается между двумя половинами аксонемы, определяемыми линией, которая проходит между центральной парой и разделяет мост и дуплет 1 пополам (рис. 2). Эта гипотеза представляет собой модель точки переключения ресничек (см. Также Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
Фиксация метахронной волны позволяет проследить развитие изгиба с разрешением электронного микроскопа. Если прикрепление спиц или междуплетов к центральной паре хотя бы частично отвечает за изгиб, как предполагали эксперименты Саммерса и Гиббонса (1971), скольжение должно вызывать смещение любой прикрепленной спицы или звена на величину скольжения. Когда изгиб происходит в основании ресничек, степень скольжения увеличивается по мере развития изгиба, и спицы наклоняются, но ни спицы, ни звенья не растягиваются непрерывно, поскольку скольжение продолжает увеличиваться; вместо этого, по мере роста изгиба вдоль аксонемы вовлекается больше спиц.
Когда изгиб происходит в основании ресничек, степень скольжения увеличивается по мере развития изгиба, и спицы наклоняются, но ни спицы, ни звенья не растягиваются непрерывно, поскольку скольжение продолжает увеличиваться; вместо этого, по мере роста изгиба вдоль аксонемы вовлекается больше спиц.
Уорнер и Сатир (1974) рассмотрели, как наклон спицы может изменяться по мере того, как изгиб увеличивается вдоль аксонемы. В пределах изгиба по мере его роста все спицы каждой группы повторений 96-нм спиц наклоняются максимум на 33 ° от нормального.Поскольку углы спиц не лежат на радиусах, проходящих через центр кривизны изгиба, и поскольку скольжение накапливается в изгибе, спицы должны быть прикреплены к выступам оболочки вокруг центральной пары, когда происходит изгиб. Спицы выровнены с двумя рядами выступов вдоль каждой из центральных микротрубочек, которые образуют центральную оболочку. Выступы повторяются и образуют нониус с радиальными спицами в точном соотношении шести повторений выступов к одному повторению группы спиц. По мере развития базального изгиба количественно добавляются новые группы дистальных спиц.
По мере развития базального изгиба количественно добавляются новые группы дистальных спиц.
Наклон спицы во внешней дублетной микротрубочке с длиной повторения 96 нм остается постоянным по мере того, как изгиб увеличивается до максимальной кривизны, затем, когда изгиб проходит, спицы отсоединяются по мере выпрямления аксонемы. В прямых участках аксонемы, проксимальных или дистальных по отношению к изгибу, относительное положение групп спиц между любыми двумя дублетами остается постоянным по длине этой области, что означает, что скольжение происходит только в областях изгиба. В прямых участках аксонемы почти все спицы каждой группы нормальны (90 °) как по отношению к их исходному дублету, так и к центральным микротрубочкам.Функционально все эти спицы следует рассматривать как отсоединенные от выступов оболочки. Наблюдаемые радиальные конфигурации спиц сильно указывают на то, что существует точный цикл отсоединения спиц — присоединения к центральной оболочке, который, как мы заключаем, составляет основную часть механизма преобразования активного междублетного скольжения в локальный изгиб.
Хотя изгибание требует активности dynein руки, локус активности руки вдоль аксонемы все еще неизвестен (см. Ниже и Lin et al. 2014).Может случиться так, что активность руки вдоль дуплета происходит только в положении сгибания. Также неизвестно, сколько циклов активности динеинового плеча необходимо для получения 96-нм единицы изгиба на одном дублете. In vitro , единственный динеин способен вызывать скольжение, но динеины внешнего плеча аксонемы имеют низкое соотношение фаза нагрузки: цикл, что означает, что для плавной непрерывной подвижности во время развития изгиба должны быть задействованы множественные динеины, действующие в противофазе. (В этой статье мы не останавливались на цикле генерации силы динеинов; отличные обзоры см. В Sakakibara and Oiwa 2011, Kikkawa 2013, and Roberts et al.2013). Разумным предположением является то, что во время развития изгиба каждый динеин — как внутренний, так и внешний — в группе спиц на активном дублете потенциально циклически повторяется один раз.
Shingyoji и его коллеги (1977) провели эксперимент с освещением. Они прикрепили демембранную прямую аксонему морского ежа к микроигле и ионтофоретически добавили небольшое количество АТФ в середину аксонемы. Поскольку аксонема была прямой и ее концы были жесткими, при активации динеиновых плеч они давали равные и противоположные изгибы.Предположительно, согласно гипотезе точки переключения, активность вдоль половины аксонемы (дублеты 1–5; рис. 2) в одной области уравновешена, противодействуя активности противоположной половины (дублеты 6–9 и 1; рис. 2) далее вдоль аксонемы. . Изгибание приводит к сопротивлению скольжению, а механическое напряжение, создаваемое действием руки на аксонему в одном положении, может передавать напряжение вдоль аксонемы, вероятно, через спицы и центральные парные соединения, чтобы активировать уравновешивающую активность руки в дублетах в более отдаленном положении — именно то, что необходимо для волновой прогрессии в гипотезе точки переключения.Модель управления кривизной Брокоу (1985) представляет собой упрощенный вид трансмиссии, который еще предстоит полностью уточнить. При нормальной перистальтике ресничек контроль такой передачи очень сложен, что позволяет большому количеству изгибных волн разной величины и времени распространяться по аксонеме. Более того, передача активности руки д. Происходить с довольно высокими скоростями, чтобы позволить завершить полный цикл биений вдоль аксонемы длиной 10-15 мкм менее чем за 10 миллисекунд в некоторых случаях.Дополнительные вопросы о колебании и изгибе ресничек обсуждаются в прекрасных недавних обзорах Brokaw (2009) и Lindemann and Lesich (2010).
При нормальной перистальтике ресничек контроль такой передачи очень сложен, что позволяет большому количеству изгибных волн разной величины и времени распространяться по аксонеме. Более того, передача активности руки д. Происходить с довольно высокими скоростями, чтобы позволить завершить полный цикл биений вдоль аксонемы длиной 10-15 мкм менее чем за 10 миллисекунд в некоторых случаях.Дополнительные вопросы о колебании и изгибе ресничек обсуждаются в прекрасных недавних обзорах Brokaw (2009) и Lindemann and Lesich (2010).
Структурная и биохимическая сложность проливает свет на нерешенные проблемы модели точки переключения
Модель точки переключения была разработана в то время, когда разрешение электронного микроскопа и биохимия аксонем были в зачаточном состоянии. Предполагалось, что динеины внешнего и внутреннего плеча представляют собой зеркально отображаемые идентичные молекулы, что оболочка вокруг центральной пары функционально симметрична, так что микротрубочки центральной пары по существу идентичны, и что биохимия аксонем была по существу простой. Ни одно из этих предположений не подтвердилось. Чтобы расширить модель для учета подвижности, потребовались новые структурные, биохимические и генетические подходы.
Ни одно из этих предположений не подтвердилось. Чтобы расширить модель для учета подвижности, потребовались новые структурные, биохимические и генетические подходы.
Важно отметить, что асимметрия в организации центральных пар обязательно означает, что в любой момент цикла изгиба отдельные проекции центральных пар взаимодействуют с радиальными спицами из единственной уникальной внешней дублетной микротрубочки (Smith EF and Yang 2004). Следовательно, одна из моделей состоит в том, что центральная пара контролирует форму волны в ресничках путем избирательной регуляции активности динеина в любой точке цикла биений на единственной внешней дублетной микротрубочке или их подмножестве.Более того, помимо асимметрии аппарата центральной пары, общей для подвижных ресничек, у многих организмов, включая Chlamydomonas , центральная пара вращается по мере изгиба ресничек (Smith EF and Yang 2004). Следовательно, в этих случаях центральная пара рассматривается как распределитель, и с вращением во время цикла биений ресничек, центральная пара, как полагают, систематически взаимодействует с разными радиальными спицами, выступающими из разных внешних дублетных микротрубочек. Предположительно, это приводит к систематической активации динеиновых моторов на разных фазах биения.Однако точная роль вращения центральной пары в ресничках у некоторых организмов не решена (Mitchell and Nakatsugawa 2004).
Предположительно, это приводит к систематической активации динеиновых моторов на разных фазах биения.Однако точная роль вращения центральной пары в ресничках у некоторых организмов не решена (Mitchell and Nakatsugawa 2004).
Асимметрия в аппарате центральной пары также подразумевает роль в избирательном контроле прямого и обратного изгиба и точки переключения, описанной выше и необходимой для чередующихся изгибов ресничек. Например, при сравнении движения жгутиков Chlamydomonas дикого типа и мутанта , Brokaw с коллегами (1982) определили, что центральная пара-радиальная система спиц важна для преобразования симметричного прямого и обратного изгибов в асимметричные изгибы, необходимые для эффективное сгибательное движение ресничек и нормальные физиологические силы.Более конкретно, одна из функций радиальных спиц, по-видимому, заключается в относительном подавлении развития обратного изгиба по сравнению с кривизной прямого изгиба (Brokaw et al. 1982).
Мутация Hydin у Chlamydomonas и у мышей выявила роль системы центральная пара-радиальная спица в контроле переключения между прямым и обратным изгибами (Lechtreck and Witman 2007, Lechtreck et al. 2008). Примечательно, что мутация в гене HYDIN приводит к большому дефекту изгиба ресничек, но структурный анализ выявляет только очень маленький дефект в структуре аксонемы.И у Chlamydomonas , и у мышей проекция одной центральной пары является единственным отсутствующим структурным компонентом мутантных аксонем. Как у Chlamydomonas , так и у мыши мутация Hydin приводит к относительной нерешительности или срыву между прямым и обратным изгибами. Следовательно, самая простая интерпретация состоит в том, что мутация hydin влияет на механизм точки переключения, необходимый для переключения между прямым и обратным изгибами (см. Комментарий Smith EF 2007).В совокупности данные показывают, что центральная пара — система радиальных спиц играет центральную роль в переключении между прямым и обратным изгибами и в управлении ими — то есть, какие дублеты имеют активные плечи, когда изгиб формируется и прогрессирует, — и в точном контроле за изгибом.
2008). Примечательно, что мутация в гене HYDIN приводит к большому дефекту изгиба ресничек, но структурный анализ выявляет только очень маленький дефект в структуре аксонемы.И у Chlamydomonas , и у мышей проекция одной центральной пары является единственным отсутствующим структурным компонентом мутантных аксонем. Как у Chlamydomonas , так и у мыши мутация Hydin приводит к относительной нерешительности или срыву между прямым и обратным изгибами. Следовательно, самая простая интерпретация состоит в том, что мутация hydin влияет на механизм точки переключения, необходимый для переключения между прямым и обратным изгибами (см. Комментарий Smith EF 2007).В совокупности данные показывают, что центральная пара — система радиальных спиц играет центральную роль в переключении между прямым и обратным изгибами и в управлении ими — то есть, какие дублеты имеют активные плечи, когда изгиб формируется и прогрессирует, — и в точном контроле за изгибом. кривизна в каждом изгибе. Однако эти модели требуют дальнейшего прямого тестирования.
кривизна в каждом изгибе. Однако эти модели требуют дальнейшего прямого тестирования.
Мутанты Chlamydomonas были особенно информативными, давая представление о составе и структуре аксонемы, регуляции подвижности, роли различных двигателей динеина, а также сборке и регуляции длины ресничек (Avasthi and Marshall 2012).Здесь мы сосредотачиваемся на том, как Chlamydomonas способствовал нашему пониманию механизма и регуляции изгиба ресничек и, в частности, как он способствовал пониманию общих ролей внешнего и внутреннего динеина в контроле частоты сердечных сокращений и формы волны. .
Первоначально и чаще всего аксонемные динеины называются внешними и внутренними динеиновыми ветвями, но на самом деле существует множество различных консервативных динеиновых моторов, которые сильно локализованы в аксонеме и служат особым целям для контроля над ней. движение (Кинг и Камия 2009).Наиболее изученным аксонемным динеином является внешнее плечо динеина. Внешние динеиновые ветви очень сложны с биохимической точки зрения: каждое состоит из двух или трех различных динеиновых АТФаз и по крайней мере 16 различных субъединиц. Они собираются индивидуально в ряды на дублетах с номерами 2–9 в аксонемах Chlamydomonas и повторяются с регулярным периодом 24 нм вдоль каждого внешнего дублета (см. Рис. 4; Гуденаф и Хойзер 1982). Вдоль большей части аксонемы каждое внешнее плечо динеина структурно — и, обычно, биохимически — идентично своему соседу.Мутации в генах, которые кодируют структурные белки внешнего плеча динеина или связанные факторы, участвующие в нацеливании и сборке внешнего плеча динеина, могут привести к неспособности собрать всю или часть внешнего плеча динеина и, как следствие, к нарушению подвижности (King и Камия 2009).
движение (Кинг и Камия 2009).Наиболее изученным аксонемным динеином является внешнее плечо динеина. Внешние динеиновые ветви очень сложны с биохимической точки зрения: каждое состоит из двух или трех различных динеиновых АТФаз и по крайней мере 16 различных субъединиц. Они собираются индивидуально в ряды на дублетах с номерами 2–9 в аксонемах Chlamydomonas и повторяются с регулярным периодом 24 нм вдоль каждого внешнего дублета (см. Рис. 4; Гуденаф и Хойзер 1982). Вдоль большей части аксонемы каждое внешнее плечо динеина структурно — и, обычно, биохимически — идентично своему соседу.Мутации в генах, которые кодируют структурные белки внешнего плеча динеина или связанные факторы, участвующие в нацеливании и сборке внешнего плеча динеина, могут привести к неспособности собрать всю или часть внешнего плеча динеина и, как следствие, к нарушению подвижности (King и Камия 2009).
Рисунок 4.
Ультраструктура аксонемы Chlamydomonas. Срезы криоэлектронной томографии показывают (а) продольный, (б) трехмерный вид и (в) поперечный разрез аксонемы Chlamydomonas. Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации. Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые аксонемные структуры: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA).Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации. Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые аксонемные структуры: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA).Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
Рисунок 4.
Ультраструктура аксонемы Chlamydomonas. Срезы криоэлектронной томографии показывают (а) продольный, (б) трехмерный вид и (в) поперечный разрез аксонемы Chlamydomonas. Красные прямоугольники выделяют одну 96-нанометровую (нм) аксонемную повторяющуюся единицу на каждом изображении. (d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации.Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые аксонемные структуры: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA). Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
(d, e) Визуализация изоповерхности и (f, g) упрощенная схема показывают усредненный аксонемный повтор 96 нм в (d, f) продольной и (e, g) поперечной ориентации.Поперечный срез берут рядом с радиальной спицей 2, если смотреть от проксимального до дистального конца. Выделены ключевые аксонемные структуры: A- и B-канальцы (A t , B t ), регуляторный комплекс нексин-динеин (N-DRC), радиальные спицы (RS1, RS2), кальмодулин и связанные спицы. комплекс (CSC), а также внутренние и внешние динеиновые рукава (IA, OA). Динеины внутреннего плеча включают комплекс I1 (динеин f α и β) и динеины a – g. Источник: адаптировано с разрешения Heuser и коллег (2012a, 2012b).
В соответствии с новаторскими исследованиями Гиббонса Б.Х. и Гиббонса (1973), наиболее заметным последствием сбоя сборки внешнего плеча динеина является снижение частоты биений ресничек (Brokaw and Kamiya 1987, Brokaw 1994, Kamiya 2002). Внешнее плечо динеина и частота биений ресничек могут регулироваться фосфорилированием и изменениями кальция (Christensen et al. 2001, King and Kamiya 2009, King 2010). У инфузорий Paramecium и Tetrahymena увеличение цАМФ вокруг аксонемы вызывает более быстрое плавание за счет фосфорилирования небольшого белка, связанного с внешним динеиновым плечом (Christensen et al.2001). In vitro , фосфорилированное внешнее плечо динеина вызывает более быстрое скольжение микротрубочек, что означает, что для получения такого же количества скольжения в течение цикла биений требуется меньше времени, то есть частота биений выше. Разнообразные доказательства также указывают на то, что внешние динеиновые рукава реагируют на механическое возмущение аксонемы (Hayashibe et al. 1997). Механическая обратная связь, вероятно, важна для переключения между активным и неактивным состояниями и необходима для прямого и обратного изгиба.В соответствии с управлением с механической обратной связью, как обсуждалось выше (Shingyoji et al. 1977), изгиб аксонемы может активировать управляемое динеином скольжение микротрубочек (Morita and Shingyoji 2004, Hayashi and Shingyoji 2008).
2001, King and Kamiya 2009, King 2010). У инфузорий Paramecium и Tetrahymena увеличение цАМФ вокруг аксонемы вызывает более быстрое плавание за счет фосфорилирования небольшого белка, связанного с внешним динеиновым плечом (Christensen et al.2001). In vitro , фосфорилированное внешнее плечо динеина вызывает более быстрое скольжение микротрубочек, что означает, что для получения такого же количества скольжения в течение цикла биений требуется меньше времени, то есть частота биений выше. Разнообразные доказательства также указывают на то, что внешние динеиновые рукава реагируют на механическое возмущение аксонемы (Hayashibe et al. 1997). Механическая обратная связь, вероятно, важна для переключения между активным и неактивным состояниями и необходима для прямого и обратного изгиба.В соответствии с управлением с механической обратной связью, как обсуждалось выше (Shingyoji et al. 1977), изгиб аксонемы может активировать управляемое динеином скольжение микротрубочек (Morita and Shingyoji 2004, Hayashi and Shingyoji 2008). Кроме того, эти данные могут также косвенно относиться к важной модели механического контроля изгиба, называемой геометрической гипотезой сцепления , которая предусматривает изменения междублетных расстояний, соответствующих активности руки и производству изгибов, связанных с искажением аксонемы во время изгиба (для полное обсуждение см. Lindemann 2011).Предполагаемое искажение аксонемы во время изгиба было обнаружено с помощью электронной микроскопии (Lindemann and Mitchell 2007).
Кроме того, эти данные могут также косвенно относиться к важной модели механического контроля изгиба, называемой геометрической гипотезой сцепления , которая предусматривает изменения междублетных расстояний, соответствующих активности руки и производству изгибов, связанных с искажением аксонемы во время изгиба (для полное обсуждение см. Lindemann 2011).Предполагаемое искажение аксонемы во время изгиба было обнаружено с помощью электронной микроскопии (Lindemann and Mitchell 2007).
Внутренние рукава динеина намного сложнее, чем внешние рукава динеина (обзоры см. В Kamiya 2002, King and Kamiya 2009). Вдоль аксонемного 96-нм повтора на дублетной микротрубочке (рисунок 4, который имеет четыре идентичных внешних плеча динеина) внутренние плечи динеина включают по крайней мере семь различных динеинов, которые различаются по составу и расположению (см. Bui et al.2012). Внутренние плечи динеина были охарактеризованы посредством биохимического фракционирования компонентов динеина и с помощью электронной микроскопии аксонем, отсутствующих подмножества внутренних плеч динеина (King and Kamiya 2009). В аксонемах Chlamydomonas , в которых отсутствует подмножество внутренних динеиновых плеч, изменяется форма волны цилиарной системы, нарушается фототаксис и замедляется скорость плавания клеток. Прямой анализ биения ресничек с помощью высокоскоростной видеозаписи подтвердил, что нарушение сборки любого из отдельных внутренних плечей динеина приводит к изменению формы волны ресничек, параметра, который важен для эффективного биения ресничек и физиологии (Brokaw and Kamiya 1987). .
В аксонемах Chlamydomonas , в которых отсутствует подмножество внутренних динеиновых плеч, изменяется форма волны цилиарной системы, нарушается фототаксис и замедляется скорость плавания клеток. Прямой анализ биения ресничек с помощью высокоскоростной видеозаписи подтвердил, что нарушение сборки любого из отдельных внутренних плечей динеина приводит к изменению формы волны ресничек, параметра, который важен для эффективного биения ресничек и физиологии (Brokaw and Kamiya 1987). .
Точная роль каждого внутреннего плеча динеина еще не изучена. Однако одно из двуглавых внутренних плечей динеина, называемое динеином I1 , считается особенно важным для контроля изгиба аксонемы. Активность I1 динеина регулируется киназами и фосфатазами, локализованными в аксонеме (см. Обзор Wirschell et al. 2011). Изменения активности I1 можно измерить по изменениям скорости скольжения микротрубочек, но пока не известно, как изменения скорости соответствуют изменениям в изгибе.Одна возможность состоит в том, что увеличение скорости скольжения, производимой внутренними плечами динеина без изменения частоты биений, будет соответствовать увеличению величины изгиба. Кроме того, I1 динеин может регулировать изгиб посредством контроля активности др. Динеинов, включая внешние динеиновые плечи и одноголовые внутренние динеиновые плечи (Kotani et al. 2007, Yamamoto et al. 2013).
Кроме того, I1 динеин может регулировать изгиб посредством контроля активности др. Динеинов, включая внешние динеиновые плечи и одноголовые внутренние динеиновые плечи (Kotani et al. 2007, Yamamoto et al. 2013).
В целом, мы еще не знаем функцию каждого из аксонемных одноголовых динеинов. Однако анализ мутанта под названием ida9 показал, что одно из внутренних плеч динеина, динеин c, необходимо для движения ресничек в вязкой жидкости (Yagi et al.2005). Кроме того, мощный скрининг выявил новые мутанты, которые регулируют активность внутреннего плеча динеина (Kamiya et al. 1991, Kamiya 2002). Напр., Скрининг выявил ферменты, ответственные за полиглутамилирование, и что эта посттрансляционная модификация тубулина является критической для активности в подмножестве внутренних плеч динеина (Kubo et al. 2010).
Центральная пара и радиальные спицы необходимы для нормальной перистальтики ресничек и управления динеиновыми моторами
У Chlamydomonas нарушение сборки центральной пары или радиальных спиц приводит либо к цилиарному параличу (Witman et al. 1978, Smith EF and Yang 2004) или в сильно измененном и непродуктивном изгибающем движении. Вместе аппарат центральной пары и радиальные спицы действуют посредством как механической, так и химической передачи сигналов, чтобы в конечном итоге контролировать активность аксонемального динеина (Smith EF and Yang 2004). Механизм взаимодействия между центральной парой и радиальной спицей был подтвержден в недавних экспериментальных исследованиях с участием Chlamydomonas (Oda et al. 2014). Они показали, что добавление неспецифических белков к головке лучевой спицы могло подавить паралич парализованного мутанта центральной пары, у которого отсутствовала часть выступов центральной пары.Простейшая интерпретация этого результата состоит в том, что добавленные белки обеспечивают физическое взаимодействие между головкой спицы и выступами, которое требуется для активации аксонемных динеинов.
1978, Smith EF and Yang 2004) или в сильно измененном и непродуктивном изгибающем движении. Вместе аппарат центральной пары и радиальные спицы действуют посредством как механической, так и химической передачи сигналов, чтобы в конечном итоге контролировать активность аксонемального динеина (Smith EF and Yang 2004). Механизм взаимодействия между центральной парой и радиальной спицей был подтвержден в недавних экспериментальных исследованиях с участием Chlamydomonas (Oda et al. 2014). Они показали, что добавление неспецифических белков к головке лучевой спицы могло подавить паралич парализованного мутанта центральной пары, у которого отсутствовала часть выступов центральной пары.Простейшая интерпретация этого результата состоит в том, что добавленные белки обеспечивают физическое взаимодействие между головкой спицы и выступами, которое требуется для активации аксонемных динеинов.
Передача сигналов от центральной пары и радиальных спиц к двигателям dynein
Анализ мутантов Chlamydomonas также выявил консервативные компоненты аксонемы, которые передают сигналы от центральной пары и структур радиальных спиц к динеиновым моторам. Наиболее примечательными являются комплекс, связанный с кальмодулином и спицами (CSC; Dymek et al. 2011, Heuser et al. 2012a) и DRC; см. следующий раздел для получения информации из недавних анализов криоэлектронной томографии (крио-ET) аксонемы (обзоры см. Heuser et al. 2009, Porter 2012) и комплекса динеина Mia-I1 (Yamamoto et al. 2013). CSC и DRC играют роль в управлении динеиновыми двигателями: они оба идеально расположены для соединения радиальных спиц с внешними дублетами и динеиновыми двигателями.В настоящее время важной целью является определение того, как кальций и комплексы кальмодулина, расположенные в центральном аппарате и CSC, управляют аксонемными динеинами. Динеин I1 и связанный с ним комплекс Mia (Yamamoto et al. 2013) также могут играть роль, сходную с ролью DRC, в регуляции или устойчивости к управляемому динеином скольжению микротрубочек и в контроле изгиба аксонемы.
Наиболее примечательными являются комплекс, связанный с кальмодулином и спицами (CSC; Dymek et al. 2011, Heuser et al. 2012a) и DRC; см. следующий раздел для получения информации из недавних анализов криоэлектронной томографии (крио-ET) аксонемы (обзоры см. Heuser et al. 2009, Porter 2012) и комплекса динеина Mia-I1 (Yamamoto et al. 2013). CSC и DRC играют роль в управлении динеиновыми двигателями: они оба идеально расположены для соединения радиальных спиц с внешними дублетами и динеиновыми двигателями.В настоящее время важной целью является определение того, как кальций и комплексы кальмодулина, расположенные в центральном аппарате и CSC, управляют аксонемными динеинами. Динеин I1 и связанный с ним комплекс Mia (Yamamoto et al. 2013) также могут играть роль, сходную с ролью DRC, в регуляции или устойчивости к управляемому динеином скольжению микротрубочек и в контроле изгиба аксонемы.
Супрессорные мутации в
Chlamydomonas и DRC Основное открытие для понимания контроля аксонемных динеинов стало результатом классических генетических исследований Huang et al. (1982).Генетический скрининг выявил новые гены, которые подавляли паралич (то есть спасательную подвижность) у мутантов с радиальными спицами или центральной парой, так что мутанты снова становились подвижными. Супрессорные мутанты включали новые мутации в моторных тяжелых цепях динеина, которые восстанавливали движение без восстановления радиальной спицы и без дефектов центральной пары. Эти результаты показывают, что в отсутствие радиальных спиц и центральной пары двигательная активность динеина подавляется по всей аксонеме, но она может быть восстановлена любым молекулярным изменением, которое позволяет динеину становиться активным без участия пары спица-центральная часть. система.
(1982).Генетический скрининг выявил новые гены, которые подавляли паралич (то есть спасательную подвижность) у мутантов с радиальными спицами или центральной парой, так что мутанты снова становились подвижными. Супрессорные мутанты включали новые мутации в моторных тяжелых цепях динеина, которые восстанавливали движение без восстановления радиальной спицы и без дефектов центральной пары. Эти результаты показывают, что в отсутствие радиальных спиц и центральной пары двигательная активность динеина подавляется по всей аксонеме, но она может быть восстановлена любым молекулярным изменением, которое позволяет динеину становиться активным без участия пары спица-центральная часть. система.
Тот же самый скрининг генетических супрессоров (Huang et al. 1982) выявил другой регуляторный комплекс, который Piperno в 1994 назвал DRC. Последующие исследования дополнительно определили белковые субъединицы DRC (для обзора см. Porter 2012). Важным недавним достижением, возможным благодаря увеличенному разрешению крио-ET, является открытие того, что DRC также является междублетным звеном нексина. Поэтому структура DRC теперь называется регуляторным комплексом нексин-динеин (N-DRC; Heuser et al.2009 г.). Недавние исследования также показали биохимически-структурное взаимодействие между N-DRC и промежуточными цепями внешнего плеча динеина, физически связывающими внешнее плечо динеина и N-DRC (Oda et al. 2013). Следовательно, N-DRC, по-видимому, играет роль в регуляции моторов динеина и служит междублетным звеном, так что он, вероятно, выравнивает внешние дублеты для обеспечения эффективных взаимодействий между моторами динеина и B-канальцем соседней микротрубочки дублета (Bower и др., 2013).Чтобы приспособиться к скольжению дублетных микротрубочек, все междублетные связи N-DRC-внешнее плечо динеина на скользящем дублете должны в какой-то момент разрывать их связь с соседней B-микротрубочкой (Holwill and Satir 1990). В окончательном понимании модели точки переключения будет важно знать способ, которым регулируются связи N-DRC, и время разрыва и реформации для активных и пассивно скользящих половин аксонемы.
Поэтому структура DRC теперь называется регуляторным комплексом нексин-динеин (N-DRC; Heuser et al.2009 г.). Недавние исследования также показали биохимически-структурное взаимодействие между N-DRC и промежуточными цепями внешнего плеча динеина, физически связывающими внешнее плечо динеина и N-DRC (Oda et al. 2013). Следовательно, N-DRC, по-видимому, играет роль в регуляции моторов динеина и служит междублетным звеном, так что он, вероятно, выравнивает внешние дублеты для обеспечения эффективных взаимодействий между моторами динеина и B-канальцем соседней микротрубочки дублета (Bower и др., 2013).Чтобы приспособиться к скольжению дублетных микротрубочек, все междублетные связи N-DRC-внешнее плечо динеина на скользящем дублете должны в какой-то момент разрывать их связь с соседней B-микротрубочкой (Holwill and Satir 1990). В окончательном понимании модели точки переключения будет важно знать способ, которым регулируются связи N-DRC, и время разрыва и реформации для активных и пассивно скользящих половин аксонемы.
Анализ cryo-ET
Новый уровень разрешения аксонемы был достигнут с помощью крио-ЭТ.Эти данные уже оказали большое влияние на понимание движения ресничек. Была рассмотрена базовая методология (например, Nicastro 2009, Bui and Ishikawa 2013) и описаны методы, которые теперь объединяют структурную локализацию с крио-ET (Oda and Kikkawa 2013). Вкратце, процесс включает в себя быстрое замораживание живых клеток или изолированных аксонем с последующей электронной микроскопией с низкой дозой облучения структур в замороженном нативном состоянии, сбор данных, компьютерную томографию и усреднение изображений, которые позволяют получать изображения с высоким разрешением в три измерения.В сочетании с очень информативными структурными мутантами в Chlamydomonas появилась картина аксонемной структуры с очень высоким разрешением. Новая структурная информация обобщена на рисунке 4, включая примеры электронных томограмм (рисунки 4a-4c), усредненные значения и трехмерные изображения изоповерхности (рисунки 4d, 4e), а также сводные схематические диаграммы одиночной дублетной микротрубочки в продольном и поперечном сечениях (рисунок 4е, 4ж).
Подход крио-ЭТ предлагает множество преимуществ: высокое разрешение, приближение и превышение 3 нм, обнаружение задержанной субструктуры каждого аксонемного компонента; первозданная сохранность за счет быстрого замораживания; и трехмерная визуализация структур в интактных органеллах.Примечательные особенности на рисунке 4 включают определение продольного 96-нм аксонемного повтора (рисунки 4 D и E), который, вероятно, является основной единицей активности вдоль аксонемы и субструктуры внешних плеч динеина, включая разрешение глобулярные моторные домены (рис. 4d – 4g). Также показаны расположение и субструктура каждого внутреннего плеча динеина (I1 / f динеин и динеины a – g; Bui et al. 2012, Heuser et al. 2012b, Lin et al. 2014), N-DRC (Heuser et al. 2009), CSC (Dymek et al.2011 г., Heuser et al. 2012a) и радиальные спицы (Pigino et al. 2011, Barber et al. 2012, Oda et al. 2014). Кроме того, крио-ЭТ выявила субструктуру аппарата центральной пары (Carbajal-Gonzalez et al. 2013, Oda et al. 2014).
2014).
За относительно короткое время эти структурные достижения внесли новое понимание структурных основ изгиба ресничек. Например, крио-ET также выявил физические связи между внешними плечами динеина, внутренними плечами динеина и N-DRC, которые могут объяснить координацию активности между структурами (например,г., Oda et al. 2013). Этот результат начинает отвечать на главный вопрос — вопрос о том, как координируется активность внешних и внутренних рукавов динеина (Kamiya 2002). Наряду с другими биофизическими и структурными исследованиями крио-ЭТ также выявило структурную основу силового удара аксонемных динеинов (Lin et al. 2014). Обещание этих новых исследований состоит в том, что эти структурные достижения будут определять структурные изменения, связанные с изгибом ресничек, и будут непосредственно тестировать модели для образования изгиба ресничек и распространения изгиба.Например, структурный анализ быстро замороженных аксонем живых сперматозоидов морского ежа показывает, что структурное состояние динеиновых моторов на дублете, расположенном на внутренней стороне изгиба, отличается от состояния динеинов, расположенных на дублетах на противоположной стороне отгиба. axoneme, на внешней стороне изгиба (см. дополнительную фигуру 1 у Lin et al. 2014). Эти данные, по-видимому, подтверждают модель точки переключения для изгиба аксонемы, возможно, раскрывая «включенные» и «выключенные» состояния динеинов (см. Brokaw, 2009).Как и ожидалось, использование крио-ЭТ и консервация путем замораживания, дальнейший анализ аксонем сперматозоидов морского ежа, мутантов Chlamydomonas и ресничек, сохраненных на стадиях метахрональных биений, предоставит исчерпывающую картину структурных изменений аксонемы и динеина, связанных с инициированием изгиба и распространение изгиба.
axoneme, на внешней стороне изгиба (см. дополнительную фигуру 1 у Lin et al. 2014). Эти данные, по-видимому, подтверждают модель точки переключения для изгиба аксонемы, возможно, раскрывая «включенные» и «выключенные» состояния динеинов (см. Brokaw, 2009).Как и ожидалось, использование крио-ЭТ и консервация путем замораживания, дальнейший анализ аксонем сперматозоидов морского ежа, мутантов Chlamydomonas и ресничек, сохраненных на стадиях метахрональных биений, предоставит исчерпывающую картину структурных изменений аксонемы и динеина, связанных с инициированием изгиба и распространение изгиба.
Выводы
Мы стоим на новом этапе понимания движения ресничек в молекулярных деталях, которые вряд ли представлялись возможными, когда были впервые предложены скользящая модель и гипотеза точки переключения.В общем, основная гипотеза, кажется, выдержала проверку временем. Есть еще важные базовые вопросы, которые необходимо решить, например, как динамические связи между структурами внутри аксонемы соответствуют точному развитию и прогрессированию изгиба (Brokaw 2009). Новые возможности включают определение функции посттрансляционных модификаций тубулина для регуляции активности динеина (например, Kubo et al. 2010). Благодаря сочетанию новых генетических и структурных методов, как было проиллюстрировано изобретательным использованием мутантов Chlamydomonas и крио-ET (e.g., Oda et al., 2014), которые привели к новому пониманию сложности механизма моторики, мы можем дать содержательный ответ на многие из оставшихся вопросов.
Новые возможности включают определение функции посттрансляционных модификаций тубулина для регуляции активности динеина (например, Kubo et al. 2010). Благодаря сочетанию новых генетических и структурных методов, как было проиллюстрировано изобретательным использованием мутантов Chlamydomonas и крио-ET (e.g., Oda et al., 2014), которые привели к новому пониманию сложности механизма моторики, мы можем дать содержательный ответ на многие из оставшихся вопросов.
Благодарности
Мы благодарны Алекса Маттейсес за помощь с цифрами. Мы также благодарны всем людям, с которыми мы работали все эти годы, нашим наставникам, студентам и нашим коллегам. Эта статья была поддержана грантом WSS от Национальных институтов здравоохранения.
Цитированная литература
.Электронная микроскопия хвоста сперматозоида; результаты, полученные с новым фиксатором
,Journal of Biophysical and Biochemical Cytology
,1959
, vol.
5
(стр.269
—278
),.Этапы цилиогенеза и регуляции длины ресничек
,Дифференциация
,2012
, т.83
(стр.S30
—S42
),,,,.Трехмерная структура радиальных спиц выявляет неоднородность и взаимодействие с динеинами в жгутиках Chlamydomonas.
,Molecular Biology of the Cell
,2012
, vol.23
(стр.111
—120
),.Механика цилиарного движения
,Биологические обзоры Кембриджского философского общества
,1974
, vol.49
(стр.85
—125
),,,,,,,.N-DRC образует консервативный биохимический комплекс, который поддерживает внешнее выравнивание дублетов и ограничивает скольжение микротрубочек в подвижных аксонемах
,Molecular Biology of the Cell
,2013
, vol.24
(стр.1134
—1152
).Несинусоидальные изгибные волны жгутиков сперматозоидов
,Journal of Experimental Biology
,1965
, vol.
43
(стр.155
—169
).Компьютерное моделирование движения жгутиков: VI. Простые модели с контролируемой кривизной определены не полностью
,Biophysical Journal
,1985
, vol.48
(стр.633
—642
).Прямые измерения скольжения между внешними дублетными микротрубочками в плавающих жгутиках сперматозоидов
,Science
,1989
, vol.243
(стр.1593
—1596
).Скольжение микротрубочек в плавающих жгутиках сперматозоидов: прямые и косвенные измерения на морских ежах и оболочках сперматозоидов
,Journal of Cell Biology
,1991
, vol.114
(стр.1201
—1215
).Контроль изгиба жгутиков: новая повестка дня, основанная на разнообразии динеина
,Подвижность клеток и цитоскелет
,1994
, vol.28
(стр.199
—204
).Размышляя о колебаниях жгутиков
,Подвижность клеток и цитоскелет
,2009
, vol.
66
(стр.425
—436
),.Характер изгиба хламидомонады жгутиков: IV. Мутанты с дефектами во внутреннем и внешнем плечах динеина указывают на различия в функции плеча динеина
,Cell Motility и Cytoskeleton
,1987
, vol.8
(стр.68
—75
),,.Анализ движения жгутиков хламидомонады : функция радиально-спицевой системы выявляется путем сравнения жгутиков дикого типа и мутантных
,Journal of Cell Biology
,1982
, vol.92
(стр.722
—732
),.Трехмерный структурный анализ жгутиков / ресничек с помощью криоэлектронной томографии
,Методы Enzymol
,2013
, vol.524
(стр.305
—323
),,,,.Полярность и асимметрия в расположении динеина и родственных структур в аксонеме Chlamydomonas
,Journal of Cell Biology
,2012
, vol.198
(стр.913
—925
),,,,,,.Консервативные структурные мотивы в центральном парном комплексе жгутиков эукариот
,Цитоскелет
,2013
, т.
70
(стр.101
—120
),,,,,,.Регуляторная легкая цепь динеина внешнего плеча ресничек у Tetrahymena thermophila
,Journal of Biological Chemistry
,2001
, vol.276
(стр.20048
—20054
),,,.CSC необходим для полной сборки радиальных спиц и подвижности ресничек дикого типа
,Molecular Biology of the Cell
,2011
, vol.22
(стр.2520
—2531
),.Исследование тонкой структуры мерцательного эпителия
,Journal of Morphology
,1954
, vol.94
(стр.221
—281
),.Движение жгутиков и активность аденозинтрифосфатазы в сперме морского ежа, экстрагированной тритоном X-100
,Journal of Cell Biology
,1972
, vol.54
(стр.75
—97
),.Влияние частичной экстракции динеиновых ветвей на движение реактивированных сперматозоидов морского ежа
,Journal of Cell Science
,1973
, vol.
13
(стр.337
—357
).Взаимосвязь между тонкой структурой и направлением биений жаберных ресничек пластинчатого моллюска
,Журнал биофизической и биохимической цитологии
,1961
, vol.11
(стр.179
—205
),.О строении жгутиков у некоторых жгутиконосцев
,Журнал биофизической и биохимической цитологии
,1960
, vol.7
(стр.697
—716
),.Динеин: белок с активностью аденозинтрифосфатазы из ресничек
,Science
,1965
, vol.149
(стр.424
—426
),.Субструктура внешнего плеча динеина
,Journal of Cell Biology
,1982
, vol.95
(стр.798
—815
). ,Цилиарное движение
,1928
Cambridge University Press
,.Механизм индуцированного колебаниями жгутика переключения активности динеина в обработанных эластазой аксонемах сперматозоидов морского ежа
,Journal of Cell Science
,2008
, vol.
121
(стр.2833
—2843
),,.Индукция временного биения парализованных жгутиков мутантов Chlamydomonas путем приложения внешней силы
,Подвижность клеток и цитоскелет
,1997
, vol.37
(стр.232
—239
),,,,.Регуляторный комплекс динеина представляет собой нексиновое звено и главный регуляторный узел в ресничках и жгутиках
,Journal of Cell Biology
,2009
, vol.187
(стр.921
—933
),,,,.CSC соединяет три основных комплекса аксонем, участвующих в регуляции динеина
,Molecular Biology of the Cell
,2012a
, vol.23
(стр.3143
—3155
),,,,,,.Криоэлектронная томография выявляет дуплет-специфические структуры и уникальные взаимодействия в динеине I1
,Proceedings of the National Academy of Science
,2012b
, vol.109
(стр.E2067
—E2076
). .Гидродинамические аспекты движения ресничек и жгутиков
,Реснички и жгутики
,1974
Academic Press
(стр.
143
—175
),.Физическая модель скольжения микротрубочек в аксонемах ресничек
,Biophysical Journal
,1990
, vol.58
(стр.905
—917
),,.Супрессорные мутации в Chlamydomonas раскрывают регуляторный механизм функции жгутиков
,Cell
,1982
, vol.28
(стр.115
—124
),.Изолированный жгутиковый аппарат Chlamydomonas : характеристика плавания вперед и изменения формы волны и обратного движения ионами кальция in vitro
,Journal of Cell Science
,1978
, vol.33
(стр.235
—253
).Функциональное разнообразие аксонемных динеинов, исследованное на мутантах Chlamydomonas
,International Review of Cytology
,2002
, vol.219
(стр.115
—155
),.Субмикромолярные уровни кальция контролируют баланс биений между двумя жгутиками в демембранных моделях Chlamydomonas
,Journal of Cell Biology
,1984
, vol.
98
(стр.97
—107
),,.Два типа жгутиковых мутантов Chlamydomonas , в которых отсутствуют различные компоненты динеина внутреннего плеча
,Journal of Cell Biology
,1991
, vol.112
(стр.441
—447
).Большие шаги к пониманию динеина
,Journal of Cell Biology
,2013
, vol.202
(стр.15
—23
).Определение механического состояния аксонемы и интеграция передачи сигналов Ca 2+ динеином внешнего плеча
,Cytoskeleton
,2010
, vol.67
(стр.207
—213
),. .Аксонемные динеины: сборка, структура и формирование силы
,Справочник по хламидомонаде: подвижность и поведение клеток
,2009
, vol.3
Academic Press
(стр.131
—208
),,,,.Механические свойства внутреннего плеча динеина-f (динеина I1) изучены с помощью тестов подвижности in vitro
,Biophysical Journal
,2007
, vol.
93
(стр.886
—894
),,,,.Полиглутамилирование тубулина регулирует подвижность аксонем путем модуляции активности динеинов внутренней части руки
,Current Biology
,2010
, vol.20
(стр.441
—445
),.Chlamydomonas reinhardtii Hydin является белком центральной пары, необходимым для подвижности жгутиков
,Journal of Cell Biology
,2007
, vol.176
(стр.473
—482
),,,,.Мутации в Hydin нарушают подвижность ресничек у мышей
,Journal of Cell Biology
,2008
, vol.180
(стр.633
—643
),,,,.Структурный механизм силового удара динеина
,Nature Cell Biology
,2014
, vol.16
(стр.479
—485
).Экспериментальные доказательства гипотезы геометрического сцепления
,Современные темы биологии развития
,2011
, vol.95
(стр.1
—31
),.
Свидетельства искривления аксонемы во время биения жгутиков Chlamydomonas
,Cell Motile and the Cytoskeleton
,2007
, vol.64
(стр.580
—589
),.Биение жгутиков и ресничек: доказанное и возможное
,Journal of Cell Science
,2010
, vol.123
(стр.519
—528
).Число фибрилл в ресничках зеленых водорослей
,Nature
,1953
, об.171
(стр.485
—486
),.Распространение изгиба приводит к вращению центральной пары у Chlamydomonas reinhardtii жгутиков
,Journal of Cell Biology
,2004
, vol.166
(стр.709
—715
),.Эффекты наложенного изгиба на скольжение микротрубочек в жгутиках сперматозоидов
,Current Biology
,2004
, vol.14
(стр.2113
—2118
).Криоэлектронная томография для изучения организации аксонем
,Методы клеточной биологии
,2009
, vol.
91
(стр.1
—39
),.Новый метод структурной маркировки с использованием криоэлектронной томографии и системы биотин-стрептавидин
,Journal of Structural Biology
,2013
, vol.183
(стр.305
—311
),,,.Идентификация внешнего-внутреннего линкера динеина как хаб-контроллера для активности аксонемального динеина
,Current Biology
,2013
, vol.23
(стр.656
—664
),,,.Передача механосигналов между центральным аппаратом и радиальными спицами контролирует активность аксонемального динеина
,Journal of Cell Biology
,2014
, vol.204
(стр.807
—819
),,,,,.Криоэлектронная томография радиальных спиц ресничек и жгутиков
,Journal of Cell Biology
,2011
, vol.195
(стр.673
—687
),,,.Мутации в «регуляторном комплексе динеина» изменяют АТФ-нечувствительные сайты связывания динеинов внутреннего плеча в аксонемах Chlamydomonas
,Journal Cell Biology
,1994
, vol.
125
(стр.1109
—1117
). .Подвижность жгутиков и регуляторный комплекс динеина
,Динеины: структура
,2012
Биология и болезнь
(стр.337
—365
),,,,.Функции и механизмы моторных белков динеина
,Nature Reviews Molecular Cell Biology
,2013
, vol.14
(стр.713
—726
),.Структура и формирование ресничек и нитей простейших рубца
,Journal of Cell Biology
,1964
, vol.20
(стр.249
—270
),.Молекулярная организация и механизм генерации силы динеина
,FEBS Journal
,2011
, vol.278
(стр.2964
—2979
),.Распространено Tetrahymena ресничек. Система для анализа расположения скользящих и аксонемных спиц
,Journal of Cell Biology
,1976
, vol.71
(стр.589
—605
),.Направление активного скольжения микротрубочек в Tetrahymena ресничках
,Proceedings of the National Academy of Science
,1977
, vol.
74
(стр.2045
—2049
).Исследования ресничек: фиксация метахрональной волны
,Journal of Cell Biology
,1963
, vol.18
(стр.345
—365
).Исследования ресничек: II. Исследование дистальной области ствола ресничек и роль нитей в подвижности
,Journal of Cell Biology
,1965
, vol.26
(стр.805
—834
).Морфологические аспекты перистальтики ресничек
,Journal of General Physiology
,1967
, vol.50
доп.
(стр.241
—258
).Исследования ресничек: 3. Дальнейшие исследования кончика ресничек и модели подвижности ресничек «скользящей нити»
,Journal of Cell Biology
,1968
, vol.39
(стр.77
—94
),.Расщепление аксонемы ресничек: значение для модели «точки переключения» активности динеинового плеча при движении ресничек
,Подвижность клеток и цитоскелет
,1989
, vol.
14
(стр.345
—358
),,.Локальная реактивация жгутиков, экстрагированных тритоном, путем ионтофоретического применения АТФ
,Nature
,1977
, vol.265
(стр.269
—270
),,.Дискретное моделирование ресничек с распределением сингулярностей: приложение к эмбриональному узлу и жидкости на поверхности дыхательных путей
,Бюллетень математической биологии
,2007
, vol.69
(стр.1477
—1510
).Hydin seek: обнаружение функции перистальтики ресничек
,Journal of Cell Biology
,2007
, vol.176
(стр.403
—404
),.Радиальные спицы и центральный аппарат: механохимические преобразователи, регулирующие подвижность жгутиков
,Подвижность клеток и цитоскелет
,2004
, vol.57
(стр.8
—17
),.Аденозинтрифосфат-индуцированное скольжение канальцев в обработанных трипсином жгутиках сперматозоидов морского ежа
,Proceedings of the National Academy of Science
,1971
, vol.
68
(стр.3092
—3096
),.Вращение и транслокация микротрубочек in vitro , индуцированная динеинами из Tetrahymena ресничек
,Cell
,1988
, vol.52
(стр.459
—469
),.Смещения микротрубочек на концах живых жгутиков
,Подвижность клеток и цитоскелет
,2002
, vol.52
(стр.151
—160
),.Структурная основа образования изгиба ресничек: изменения положения радиальных спиц, сопровождающие скольжение микротрубочек
,Journal of Cell Biology
,1974
, vol.63
(стр.35
—63
),,,,,.Регуляция подвижности ресничек: консервативные протеинкиназы и фосфатазы нацелены и закреплены в аксонеме ресничек
,Архив биохимии и биофизики
,2011
, vol.510
(стр.93
—100
),,.Chlamydomonas жгутиковых мутантов без радиальных спиц и центральных канальцев: структура, состав и функция специфических аксонемных компонентов
,Journal of Cell Biology
,1978
, vol.
76
(стр.729
—747
),,,,,,.Аксонемный динеин, особенно важный для движения жгутиков при высокой вязкости: последствия от нового мутанта Chlamydomonas , дефицитного по гену тяжелой цепи динеина DHC9
,Journal of Biological Chemistry
,2005
, vol.280
(стр.41412
—41420
),,,,,,,,,.Комплекс MIA представляет собой консервативный и новый регулятор динеина, необходимый для нормальной подвижности ресничек
,Journal of Cell Biology
,2013
, vol.201
(стр.263
—278
) © Автор (ы) 2014. Опубликовано Oxford University Press от имени Американского института биологических наук. Все права защищены. Для получения разрешений, пожалуйста, пишите на электронную почту: журналы[email protected].% PDF-1.3
%
265 0 объект>
endobj
xref
265 92
0000000016 00000 н.
0000003086 00000 н.
0000003396 00000 н. 0000003431 00000 н.
0000003586 00000 н.
0000004203 00000 н.
0000004296 00000 н.
0000004391 00000 п.
0000004486 00000 н.
0000004581 00000 п.
0000004676 00000 н.
0000004770 00000 н.
0000004866 00000 н.
0000004962 00000 н.
0000005058 00000 н.
0000005154 00000 н.
0000005250 00000 н.
0000005346 00000 п.
0000005442 00000 н.
0000005585 00000 н.
0000005705 00000 н.
0000005841 00000 н.
0000006746 00000 н.
0000007212 00000 н.
0000007839 00000 п.
0000008740 00000 н.
0000009185 00000 п.
0000009590 00000 н.
0000009821 00000 н.
0000009918 00000 н.
0000010107 00000 п.
0000010419 00000 п.
0000010579 00000 п.
0000010776 00000 п.
0000011439 00000 п.
0000014228 00000 п.
0000016948 00000 н.
0000017842 00000 п.
0000018150 00000 п.
0000018384 00000 п.
0000018494 00000 п.
0000018545 00000 п.
0000018695 00000 п.
0000018859 00000 п.
0000019081 00000 п.
0000019331 00000 п.
0000019542 00000 п.
0000019829 00000 п.
0000019892 00000 п.
0000020076 00000 п.
0000020481 00000 п.
0000003431 00000 н.
0000003586 00000 н.
0000004203 00000 н.
0000004296 00000 н.
0000004391 00000 п.
0000004486 00000 н.
0000004581 00000 п.
0000004676 00000 н.
0000004770 00000 н.
0000004866 00000 н.
0000004962 00000 н.
0000005058 00000 н.
0000005154 00000 н.
0000005250 00000 н.
0000005346 00000 п.
0000005442 00000 н.
0000005585 00000 н.
0000005705 00000 н.
0000005841 00000 н.
0000006746 00000 н.
0000007212 00000 н.
0000007839 00000 п.
0000008740 00000 н.
0000009185 00000 п.
0000009590 00000 н.
0000009821 00000 н.
0000009918 00000 н.
0000010107 00000 п.
0000010419 00000 п.
0000010579 00000 п.
0000010776 00000 п.
0000011439 00000 п.
0000014228 00000 п.
0000016948 00000 н.
0000017842 00000 п.
0000018150 00000 п.
0000018384 00000 п.
0000018494 00000 п.
0000018545 00000 п.
0000018695 00000 п.
0000018859 00000 п.
0000019081 00000 п.
0000019331 00000 п.
0000019542 00000 п.
0000019829 00000 п.
0000019892 00000 п.
0000020076 00000 п.
0000020481 00000 п. 0000021144 00000 п.
0000023997 00000 п.
0000025497 00000 п.
0000026971 00000 п.
0000028619 00000 п.
0000029152 00000 п.
0000029208 00000 п.
0000029514 00000 п.
0000029919 00000 н.
0000030582 00000 п.
0000030693 00000 п.
0000030814 00000 п.
0000032411 00000 п.
0000034702 00000 п.
0000036273 00000 п.
0000036317 00000 п.
0000036371 00000 п.
0000036548 00000 н.
0000036775 00000 п.
0000037954 00000 п.
0000038338 00000 п.
0000038526 00000 п.
0000038630 00000 п.
0000039035 00000 п.
0000050520 00000 п.
0000054783 00000 п.
0000065498 00000 п.
0000072875 00000 п.
0000073649 00000 п.
0000074054 00000 п.
0000074156 00000 п.
0000074819 00000 п.
0000075082 00000 п.
0000076658 00000 п.
0000078018 00000 п.
0000078681 00000 п.
0000080643 00000 п.
0000081890 00000 п.
0000082000 00000 п.
0000084567 00000 п.
0000002136 00000 п.
трейлер
] >>
startxref
0
%% EOF
356 0 obj> поток
xb«`c`g`p Ā
0000021144 00000 п.
0000023997 00000 п.
0000025497 00000 п.
0000026971 00000 п.
0000028619 00000 п.
0000029152 00000 п.
0000029208 00000 п.
0000029514 00000 п.
0000029919 00000 н.
0000030582 00000 п.
0000030693 00000 п.
0000030814 00000 п.
0000032411 00000 п.
0000034702 00000 п.
0000036273 00000 п.
0000036317 00000 п.
0000036371 00000 п.
0000036548 00000 н.
0000036775 00000 п.
0000037954 00000 п.
0000038338 00000 п.
0000038526 00000 п.
0000038630 00000 п.
0000039035 00000 п.
0000050520 00000 п.
0000054783 00000 п.
0000065498 00000 п.
0000072875 00000 п.
0000073649 00000 п.
0000074054 00000 п.
0000074156 00000 п.
0000074819 00000 п.
0000075082 00000 п.
0000076658 00000 п.
0000078018 00000 п.
0000078681 00000 п.
0000080643 00000 п.
0000081890 00000 п.
0000082000 00000 п.
0000084567 00000 п.
0000002136 00000 п.
трейлер
] >>
startxref
0
%% EOF
356 0 obj> поток
xb«`c`g`p Ā
[1] | Афзелиус
Б. |
[2] | Антоний Д. , Беккер-Хек А. , Заривала М.А. , Шмидтс М. , Onoufriadis А. , Форухан М. , Уилсон Р. , Тейлор-Кокс Т. , Дьюар А. , Джексон С. , Гоггин П. , Ложи N.T. , Ольбрих ЧАС. , Ясперс М. , Йориссен М. , Ли М.В. , Волк МЫ. , Дэниелс М.Л. , Никто П.Г. , Ферколь T.W. , Сагель S.D. , Розенфельд М. , Рутман А., Диксит А. , О’Каллаган С. , Лукас J.S. , Hogg С. , Скамблер П.Дж. , Emes R.D. , Uk10k , Чанг ЭМ. , Shoemark А. , Ноулз МИСТЕР. , Омран ЧАС. и Mitchison H.M. , Мутации в CCDC39 и CCDC40 являются основной причиной первичной дискинезии ресничек с дезорганизацией аксонем и отсутствием внутренних плеч динеина, Hum Mutat 34 (2013), 462–472. |
[3] | Арнал
J.F.
,
Флорес
П.
,
Рами
Дж.
,
Мюррис-Эспин
М.
,
Bremont
Ф.,
Пасто
Я.
,
Серрано
Э.
и
Дидье
А.
, Концентрация оксида азота в носу при воспалительных заболеваниях околоносовых пазух, Eur Respir J 13 (1999), 307–312. |
[4] | Остин-Цзе С. , Алебриттер Дж. , Заривала М.А. , Gilberti R.M. , Ну и дела H.Y. , Хеллман Н. , Патхак Н. , Лю Ю. , Паницци J.R. , Патель-Кинг Р.С. , Tritschler Д. , Беседка Р. , О’Тул Э. , Порат J.D. , Херд T.W. , Чаки М. , Диас К.А. , Коль С., Ловрич С. , Hwang Д.Ю. , Браун Д.А. , Schueler М. , Айрик Р. , Отто E.A. , Ли М.В. , Никто П.Г. , Карсон J.L. , Дэвис S.D. , Питтман J.E. , Ферколь T.W. , Аткинсон J.J. , Оливье К. , Сагель S.D. , Dell S.D. , Розенфельд М. , Милла C.E. , Ложи N.T. , Омран ЧАС. , Портье МНЕ. , король С.М. , Ноулз МИСТЕР. , Драммонд Я. , Хильдебрандт Ф. , Скрининг цилиопатии рыбок данио плюс мутационный анализ человека идентифицируют дефекты C21orf59 и CCDC65 как вызывающие первичную цилиарную дискинезию, Am J Hum Genet 93 (2013), 672–686. |
[5] | Авата
Дж.
,
Такада
С.
,
Standley
С.
,
Lechtreck
К.Ф.
,
Bellve
К.Д.
,
Пазур
G.J.
,
Фогарти
К.Э.
и
Витман
Г. Б.
, NPHP4 контролирует цилиарный транспорт мембранных белков и крупных растворимых белков в переходной зоне, J Cell Sci 127 (2014), 4714–4727. |
[6] | Бейкер К. и Beales П.Л. , Понимание ресничек при болезни: цилиопатии человека, Am J Med Genet C Semin Med Genet 151C (2009), 281–295. |
[7] | Барбато А. , Фришер Т. , Куехни C.E. , Снайдерс Д. , Азеведо Я. , Бактай ГРАММ. , Бартолони Л. , Эбер Э. , Escribano А. , Хаарман Э. , Hesselmar Б. , Hogg С. , Йориссен М. , Лукас Дж. , Nielsen КГ. , О’Каллаган С. , Омран ЧАС. , Похунек П. , Стрипполи М.П. и куст А. , Первичная цилиарная дискинезия: согласованное заявление о подходах к диагностике и лечению у детей, Eur Respir J 34 (2009), 1264–1276. |
[8] | Басу Б. и Брюкнер М. , Многофункциональные органеллы ресничек в центре лево-правой асимметрии позвоночных, Curr Top Dev Biol 85 (2008), 151–174. |
[9] | Беккер-Хек
А.
,
Зон
I.E.
,
Окабе
Н.
,
Поллок
А.
,
Ленхарт
К.Б.
,
Салливан-Браун
Дж.
,
МакШин
Дж. |
[10] | Бейдон Н. , Chambellan А. , Альберти С. , де Блик Дж. , Климент А. , Escudier Э. и Ле Буржуа М. , Технические и практические вопросы измерения содержания оксида азота в носу при приливном дыхании у детей, Pediatr Pulmonol 50 (2015), 1374–1382. |
[11] | Бланшон
С.
,
Legendre
М.
,
Копин
Б.
,
Duquesnoy
П.
,
Монтантин
ГРАММ.
,
Котт
Э.
,
Дастот
Ф.
,
Джинсон
Л.
,
Cachanado
М.
,
Руссо
А.
,
Папон
J.F.
,
Бейдон
Н.
,
Brouard
Дж.
,
Crestani
Б.
,
Дешильдре
А.
,
Желание
Дж.
,
Дольфус
ЧАС.
,
Leheup
Б.
,
Тамалет
А.
,
Тюмерель
С.
,
Войтек
ЯВЛЯЮСЬ.
,
Escalier
Д.
,
Косте
А. |
[12] | Боаретто Ф. , Снайдерс Д. , Сальворо С. , Spalletta А. , Mostacciuolo М.Л. , Collura М. , Cazzato С. , Girosi Д. , Сильвестри М. , Росси Г.А. , Барбато А. и Вазза ГРАММ. , Диагностика первичной цилиарной дискинезии с помощью целевой панели секвенирования следующего поколения: молекулярные и клинические данные у итальянских пациентов, J Mol Diagn 18 (2016), 912–922. |
[13] | Bonnefoy
С.,
Watson
СМ.
,
Kernohan
К.Д.
,
Лемос
М.
,
Hutchinson
С.
,
Поултер
J.A.
,
Crinnion
L.A.
,
ягода
Я.
,
Симмондс
Дж.
,
Васудеван
П.
,
О’Каллаган
С.
,
Hirst
Р.А.
,
Рутман
А.
,
Хуанг
Л.
,
Хартли
Т.
,
Grynspan
Д.
,
Моя
Э.
,
Ли
С.
,
Карр
Я.
,
Bonthron
D.T.
,
Леру
М.
,
Бойкотировать
К.
,
Бастин
П.
и
Шеридан
НАПРИМЕР.
, Двуаллельные мутации в LRRC56, Кодирование белка, связанного с внутрижладжеллярным транспортом, вызывают мукоцилиарный клиренс и дефекты латеральности, Am J Hum Genet 103 (2018), 727–739. |
[14] | Благо М. , Мейтс Я. , Proesmans М. , Vermeulen F.L. , Йориссен М. и Де Бок, К. , Диагностическая точность измерения оксида азота для выявления первичной цилиарной дискинезии, Eur J Clin Invest 44 (2014), 477–485. |
[15] | Благо М. , Wallmeier Дж. , Ма Л. , Ложи N.T. , Ясперс М. , Ольбрих ЧАС. , Догерти G.W. , Raidt Дж. , Вернер С., Амирав Я. , Хеврони А. , Abitbul Р. , Авиталь А. , Соферман Р. , Wessels М. , О’Каллаган С. , Чанг ЭМ. , Рутман А. , Hirst Р.А. , Моя Э. , Mitchison H.M. , Van Daele С. , Де Бок К. , Йориссен М. , Кинтнер С. , Cuppens ЧАС. и Омран ЧАС. , Мутации MCIDAS приводят к нарушению мукоцилиарного клиренса с уменьшением образования множественных подвижных ресничек, Nat Commun 5 (2014), 4418. |
[16] | Беседка
Р.,
Tritschler
Д.
,
Vanderwaal
К.
,
Перроне
C.A.
,
Мюллер
Дж.
,
Лиса
Л.
,
распродажа
W.S.
и
Портье
МНЕ.
, N-DRC формирует консервативный биохимический комплекс, который поддерживает выравнивание внешнего дублета и ограничивает скольжение микротрубочек в подвижных аксонемах, Mol Biol Cell 24 (2013), 1134–1152. |
[17] | Brokaw C.J. и Камия Р. , Особенности изгиба жгутиков хламидомонады: IV. Мутанты с дефектами во внутреннем и внешнем плечах динеина указывают на различия в функции плеча динеина, Cell Motil Cytoskeleton 8 (1987), 68–75. |
[18] | коричневый D.E. , Питтман J.E. , Ли М.В. , Fordham Л. и Дэвис S.D. , Раннее заболевание легких у детей раннего возраста с первичной цилиарной дискинезией, Pediatr Pulmonol 43 (2008), 514–516. |
[19] | Брюкнер М. , Гетеротаксия, врожденные пороки сердца и первичная цилиарная дискинезия, Circulation 115 (2007), 2793–2795. |
[20] | Будный
Б.,
Чен
W.
,
Омран
ЧАС.
,
Fliegauf
М.
,
Цшах
А.
,
Вишневская
М.
,
Дженсен
L.R.
,
Рейно
М.
,
Шойхет
S.A.
,
Бадура
М.
,
Ленцнер
С.
,
Латош-Беленска
А.
и
Ropers
H.H.
, Новый синдром X-сцепленной рецессивной умственной отсталости, включающий макроцефалию и цилиарную дисфункцию, является аллельным к орально-лицевому-цифровому синдрому типа I, Hum Genet 120 (2006), 171–178. |
[21] | Бустаманте-Марин X.M. , Инь W.N. , Sears П.Р. , Вернер МНЕ. , Brotslaw E.J. , Митчелл Б.Дж. , Jania СМ. , Земан К.Л. , Роджерс Т.Д. , сельдь L.E. , Рефаберт Л. , Томас Л. , Амселем С. , Escudier Э. , Legendre М. , Грабб Б.Р. , Ноулз МИСТЕР. , Заривала М.А. и Островский L.E. Отсутствие GAS2L2 вызывает PCD, нарушая ориентацию ресничек и мукоцилиарный зазор, Am J Hum Genet 104 (2019), 229–245. |
[22] | Кэмпбелл Р. , Лечение осложнений со стороны верхних дыхательных путей при первичной цилиарной дискинезии у детей, Curr Opin Allergy Clin Immunol 12 (2012), 32–38. |
[23] | Каспары Т. , Ларкинс C.E. и Андерсон К.В. , Градуированный ответ на Sonic Hedgehog зависит от архитектуры ресничек, Dev Cell 12 (2007), 767–778. |
[24] | Castleman
V.H.
,
Romio
Л.
,
Чодхари
Р.
,
Hirst
Р.А.
,
де Кастро
S.C. |
[25] | Чен Ю. , Чжао М. , Ван С. , Чен Дж. , Ван Ю. , Цао В. , Чжоу W., Лю Дж. , Сюй Z. , Тонг ГРАММ. и Ли Дж. , Новая роль DYX1C1, белка-шаперона как для Hsp70, так и для Hsp90, при раке груди, J Cancer Res Clin Oncol 135 (2009), 1265–1276. |
[26] | Кристенсен
S.T.
,
Педерсен
С.Ф.
,
Сатир
П.
,
Веланд
I.R.
и
Шнайдер
Л.
, Первичная ресничка координирует сигнальные пути в контроле клеточного цикла и миграции во время развития и восстановления тканей, Curr Top Dev Biol 85 (2008), 261-301. |
[27] | Коул Д.Г. , Динер D.R. , Himelblau А.Л. , Бук П.Л. , Fuster J.C. и Розенбаум J.L. , Chlamydomonas kinesin-II-зависимый внутрижладжковый транспорт (IFT): IFT частицы содержат белки, необходимые для сборки ресничек в сенсорных нейронах Caenorhabditis elegans, J Cell Biol 141 (1998), 993–1008. |
[28] | Коллинз S.A. , Бехан Л. , Харрис А. , Gove К.и Лукас J.S. Опасности широко распространенного скрининга оксида азота на предмет первичной цилиарной дискинезии, Thorax 71 (2016), 560–561. |
[29] | Коллинз S.A. , Gove К. , Уокер W. и Лукас J.S. , Назальный скрининг оксида азота на предмет первичной цилиарной дискинезии: систематический обзор и метаанализ, Eur Respir J 44 (2014), 1589–1599. |
[30] | Dalrymple
Р.А.
и
Кения
П., Руководство Европейского респираторного общества по диагностике первичной цилиарной дискинезии: обзор рекомендаций, Arch Dis Child Educ Pract. |
[31] | Damseh Н. , Quercia Н. , Румман Н. , Dell S.D. и Ким R.H. , Первичная цилиарная дискинезия: механизмы и лечение, Appl Clin Genet 10 (2017), 67–74. |
[32] | Дэниелс М.Л. и Никто П.Г. , Генетика, диагностика и будущие стратегии лечения первичной цилиарной дискинезии, Expert Opin Orphan Drugs 3 (2015), 31–44. |
[33] | Давенпорт J.R. и Йодер Б.К. , Невероятное десятилетие для первичной реснички: взгляд на когда-то забытую органеллу, F, Am J Physiol Renal Physiol 289 (2005), 1159–1169. |
[34] | Дэвис
S.D.
,
Ферколь
T.W.
,
Розенфельд
М.
,
Ли
H.S.
,
Dell
S.D.
,
Сагель
S.D.
,
Милла
С.
,
Заривала
М.А.
,
Питтман
J.E.
,
Шапиро
А.Дж.
,
Карсон
J.L.
,
Кришер
Дж.П.
,
Hazucha
M.J.
,
Купер
М.Л.
,
Ноулз
МИСТЕР.
и
Ли
М.В.
, Клинические особенности первичной цилиарной дискинезии у детей по генотипу и ультраструктурному фенотипу, Am J Respir Crit Care Med 191 (2015), 316–324. |
[35] | Дэвис S.D. , Розенфельд М. , Ли H.S. , Ферколь T.W. , Сагель S.D. , Dell S.D. , Милла С. , Питтман J.E. , Шапиро А.Дж. , Салливан К. , Nykamp K.R. , Кришер J.P. , Заривала М.А. , Ноулз М.Р. , Ли М.В. и Консорциум по генетическим заболеваниям мукоцилиарного клиренса, Первичная цилиарная дискинезия: продольное исследование заболевания легких по дефекту ультраструктуры и генотипу, Am J Respir Crit Care Med 199 (2018), 190–198. |
[36] | де Ионг RU. и Rutland Дж. , Дефекты ресничек у здоровых людей, бронхоэктазы и первичная цилиарная дискинезия, Am J Respir Crit Care Med 151 (1995), 1559–1567. |
[37] | де Санти М.М. , Гарди С. , Barlocco ГРАММ. , Canciani М. , Мастелла ГРАММ. и Лунгарелла ГРАММ. , Дыхательные клетки без ресничек при аплазии ресничек, Biol Cell 64 (1988), 67–70. |
[38] | Догерти
G. |
[39] | Дурье Б. , Duquesnoy П., Escudier Э. , Бриду ЯВЛЯЮСЬ. , Escalier Д. , Rayet Я. , Маркос Э. , Войтек ЯВЛЯЮСЬ. , Bercher J.F. и Амселем С. , Распространенный вариант в сочетании с нонсенс-мутацией в члене семейства тиоредоксинов вызывает первичную цилиарную дискинезию, Proc Natl Acad Sci U S. A 104 (2007), 3336–3341. |
[40] | Эль-Сайед
Ю. |
[41] | Элиассон Р. , Mossberg Б. , Camner П. и Афзелиус Б.А. , Синдром неподвижно-ресничного. Врожденная цилиарная аномалия как этиологический фактор хронических инфекций дыхательных путей и мужского бесплодия, N Engl J Med 297 (1977), 1–6. |
[42] | Эллерман А. и Bisgaard ЧАС. , Продольное исследование функции легких в группе первичной цилиарной дискинезии, Eur Respir J 10 (1997), 2376–2379. |
[43] | Эсснер J.J. , Воган К.Дж. , Вагнер М.К. , Табин C.J. , Йост H.J. и Брюкнер М. , Консервативная функция эмбриональных узловых ресничек, Nature 418 (2002), 37–38. |
[44] | Фальк Н. , Losl М. , Шредер Н. и Giessl А. , Специализированные реснички сенсорных систем млекопитающих, Cells 4 (2015), 500–519. |
[45] | Фассад М.Р. , Shoemark А. , Legendre М. , Hirst Р.А. , Koll Ф. , Borgne П. , Луи Б. , Даудвохра Ф. , Патель М.П. , Томас Л. , Диксон М. , Burgoyne Т. , Hayes Дж. , Николсон А.Г. , Cullup Т. , Дженкинс Л. , Карр С. , Аврора П. , Лемуллуа М. , Обюссон-Флери А. , Папон J.F. , О’Каллаган С. , Амселем, С. , Hogg С. , Escudier Э. , Тассин ЯВЛЯЮСЬ. и Mitchison H.M. , Мутации в ДНКH9 тяжелой цепи внешнего плеча динеина вызывают дефекты подвижных ресничек и обратную сторону, Am J Hum Genet 103 (2018), 984–994. |
[46] | Ферколь Т. и Ли М. , Первичная цилиарная дискинезия и респираторный дистресс у новорожденных, Semin Perinatol 30 (2006), 335–340. |
[47] | Ферколь T.W. и Ли М.В. , Цилиопатии: центральная роль ресничек в спектре детских заболеваний, J Pediatr 160 (2012), 366–371. |
[48] | Ферранте М.Я. , Зулло А. , Барра А. , Бимонте С. , Messaddeq Н. , Studer М. , Долле П. и Франко Б. , Oral-facial-digital type I protein необходим для формирования первичных ресничек и спецификации оси слева-справа, Nat Genet 38 (2006), 112–117. |
[49] | Фаукс МНЕ. и Митчелл D.R. , Роль предварительно собранных цитоплазматических комплексов в сборке субъединиц динеина жгутика, Mol Biol Cell 9 (1998), 2337–2347. |
[50] | Фроммер А., Hjeij Р. , Ложи N.T. , Эдельбуш С. , Янке С. , Raidt Дж. , Вернер С. , Wallmeier Дж. , Гросс-Оннебринк Дж. , Ольбрих ЧАС. , Синдрик С. , Ясперс М. , Благо М. , Мемари Ю. , Дурбин Р. , Кольб-Кокочинский А. , Зауэр С. , Мартин J.K. , Nielsen КГ. , Амирав Я. , Элиас Н. , Керем Э. , Шосеев Д. , Haeffner К. и Омран ЧАС. , Иммунофлуоресцентный анализ и диагностика первичной цилиарной дискинезии с дефектами радиальных спиц, Am J Respir Cell Mol Biol 53 (2015), 563–573. |
[51] | Гердес J.M. , Дэвис E.E. и Кацанис Н. , Первичная ресничка позвоночных в развитии, гомеостазе и заболевании, Cell 137 (2009), 32–45. |
[52] | Гишар С. , Харрикейн M.C. , Лафит J.J. , Годар П. , Заегель М. , Tack В. , Лалау ГРАММ. и Бувань П. , Мутации гена промежуточной цепи аксонемального динеина (DNAI1) приводят к обратному развитию и первичной цилиарной дискинезии (синдром Картагенера), Am J Hum Genet 68 (2001), 1030–1035. |
[53] | Хэдфилд П.Дж. , Роу-Джонс J.M. , куст А. и Маккей ЯВЛЯЕТСЯ. , Лечение экссудативного среднего отита у детей с первичной цилиарной дискинезией, Clin Otolaryngol Allied Sci 22 (1997), 302–306. |
[54] | Halbeisen Ф.С. , Гутаки М. , Spycher B.D. , Амирав Я. , Бехан Л. , Благо М. , Hogg С. , Casaulta С. , Кроули С. , Хаарман НАПРИМЕР. , Карадаг Б., Кёрнер-Реттберг С. , Лёбингер МИСТЕР. , Мазурек ЧАС. , Морган Л. , Nielsen КГ. , Омран ЧАС. , Санта Мария Ф. , Schwerk Н. , Thouvenin ГРАММ. , Yiallouros П. , Лукас J.S. , Латцин П. и Куехни C.E. , Функция легких у пациентов с первичной цилиарной дискинезией: когортное исследование iPCD, Eur Respir J 52 (2018). |
[55] | Харрисон M.J. , Шапиро А.Дж. и Кеннеди М.П. , Врожденный порок сердца и первичная цилиарная дискинезия, Paediatr Respir Rev 18 (2016), 25–32. |
[56] | Hellinckx Дж. , Демедтс М. и Де Бок К. , Первичная цилиарная дискинезия: эволюция легочной функции, Eur J Pediatr 157 (1998), 422–426. |
[57] | Heuser Т. , Райчев М. , Krell Дж. , Портье МНЕ. и Никастро Д. , Регуляторный комплекс динеина представляет собой нексиновое звено и главный регуляторный узел в ресничках и жгутиках, J Cell Biol 187 (2009), 921–933. |
[58] | Хирокава Н. , Танака Ю. , Окада Ю. и Такеда С. , Узловой поток и создание лево-правой асимметрии, Cell 125 (2006), 33–45. |
[59] | Hoben Я. , Hjeij Р. , Ольбрих ЧАС. , Догерти G.W. , Nothe-Menchen Т. , Апреа Я. , откровенный Д. , Pennekamp П. , Дворничак Б. , Wallmeier Дж. , Raidt Дж. , Nielsen КГ. , Филипсен M.C. , Санта Мария Ф., Venditto Л. , Амирав Я. , Муссаффи ЧАС. , Prenzel Ф. , Ву К. , Бейки Z. , Шмидтс М. , Ложи N.T. и Омран ЧАС. , Мутации в C11orf70 вызывают первичную цилиарную дискинезию с рандомизацией симметрии левого / правого тела из-за дефектов внешнего и внутреннего плеч динеина, Am J Hum Genet 102 (2018), 973–984. |
[60] | Hong D.H. , Павлик Б. , Соколов М. , Strissel К.Дж. , Ян Дж. , Tulloch Б. , Райт А.Ф. , Аршавский В.Ю. и Ли Т. , Изоформы RPGR в фоторецепторах, соединяющих реснички и переходную зону подвижных ресничек, Invest Ophthalmol Vis Sci 44 (2003), 2413–2421. |
[61] | Horani А. , Броды S.L. , Ферколь Т. , Шосеев Д. , Вассерман М.Г. , Та-шма А. , Уилсон К.С. , Бейли П.В. , Амирав Я. , Коэн-Цемберкно М. , Датчер С.К. , Эльпелег О. и Керем Э. , Мутация CCDC65 вызывает первичную дискинезию ресничек с нормальной ультраструктурой и гиперкинетическими ресничками, PLoS One 8 (2013), e72299. |
[62] | Horani А. , Друлей T.E. , Заривала М.А. , Патель A.C. , Левинсон Б.Т. , Ван Арендонк L.G. , Торнтон К.С. , Джакалоне J.C. , Олби А.Дж. , Уилсон К.С. , Тернер E.H. , Никерсон Д.А. , Shendure Дж. , Бейли П.В. , Ли М.В. , Ноулз МИСТЕР. , Броды S.L. , Датчер С.К. и Ферколь T.W. , Захват всего экзома и секвенирование идентифицируют мутацию HEATR2 как причину первичной цилиарной дискинезии, Am J Hum Genet 91 (2012), 685–693. |
[63] | Horani А. и Ферколь T.W. , Первичная цилиарная дискинезия и ассоциированные сенсорные цилиопатии, Expert Rev Respir Med 10 (2016), 569–576. |
[64] | Horani А. , Ферколь T.W. , Шосеев Д. , Вассерман М.Г. , Орен Ю.С. , Керем Б. , Амирав Я. , Коэн-Цемберкно М. , Датчер С.К. , Броды S.L. , Эльпелег О. и Керем Э. , Мутация LRRC6 вызывает первичную цилиарную дискинезию с дефектами динеинового плеча, PLoS One 8 (2013), e59436. |
[65] | Horani А. , Ustione А. , Хуанг Т. , Ферт А.Л. , Сковорода Дж. , Gunsten С.П. , Хаспел J.A. , Поршень Д.В. и Броды S.L. , Установление белкового комплекса предварительной сборки ресничек во время подвижного цилиогенеза, Proc Natl Acad Sci U S. A 115 (2018), E1221 – E1228. |
[66] | Хорват Дж. , Fliegauf М. , Ольбрих ЧАС. , Kispert А. , король С.М. , Mitchison ЧАС., Заривала М.А. , Ноулз МИСТЕР. , Судбрак Р. , Фекете ГРАММ. , Neesen Дж. , Рейнхардт Р. и Омран ЧАС. , Идентификация и анализ аксона 1 легкой цепи динеина эмали у пациентов с первичной цилиарной дискинезией, Am J Respir Cell Mol Biol 33 (2005), 41–47. |
[67] | Hossain Т. , Каппельман M.D. , Перес-Атайде A.R. , Молодой G.J. , Huttner К. и Христу ЧАС. , Первичная цилиарная дискинезия как причина респираторного дистресса новорожденных: значение для неонатолога, J. Perinatol 23 (2003), 684–687. |
[68] | Инаба К. и Mizuno К. , Дисфункция сперматозоидов и цилиопатия, Reprod Med Biol 15 (2016), 77–94. |
[69] | Исикава Т. , Сакакибара ЧАС. и Оива К. , Архитектура внешних рукавов динеина, J Mol Biol 368 (2007), 1249–1258. |
[70] | Джайн Р. , Джавидан-Нежад С. , Александр-Бретт Дж. , Horani А., Cabellon M.C. , Уолтер M.J. и Броды S.L. , Сенсорные функции подвижных ресничек и их влияние на бронхоэктазы, Front Biosci (Schol Ed) 4 (2012), 1088–1098. |
[71] | Джинсон Л. , Копин Б. , Папон J.F. , Дасто-Ле Моаль Ф. , Дукесной П. , Монтантин Г. , Кадранель Дж. , Корвол Х. , Косте А. , Дезир Дж. , Суая А. , Котт Э. , Колло Н. , Тиссье С. , Луи Б. , Тамалет А. , де Блик Дж. , Клемент А. , Эскудье Э., Амселем С. и Лежандр М. , Мутации RSPh4 вызывают первичную дискинезию ресничек с дефектами центрального комплекса и почти полным отсутствием радиальных спиц, Am J Hum Genet 97 (2015), 153–162. |
[72] | Джонсон N.T. , Виллалон М. , Ройс F.H. , Жесткий Р. и Вердуго П. , Ауторегуляция частоты биений в респираторных реснитчатых клетках. Демонстрация вязкой нагрузки, Am Rev Respir Dis 144 (1991), 1091–1094. |
[73] | Картагенер М., Zur pathogenese der bronkiectasien. Bronkiectasien bei situs viscerum inversus, Beitr Klin Tuberk 82 (1933), 489–501. |
[74] | Келлетт Ф. и Роберт Н.М. , Распыленный 7% гипертонический раствор улучшает функцию легких и качество жизни при бронхоэктазах, Respir Med 105 (2011), 1831–1835. |
[75] | Кеннеди М.П. , Никто П.Г. , Ли М.В. , Заривала М.А. , Минникс С.Л. , Ноулз МИСТЕР. и Молина П.Л. , КТ высокого разрешения пациентов с первичной цилиарной дискинезией, AJR Am J Roentgenol 188 (2007), 1232–1238. |
[76] | Кеннеди М.П. , Омран ЧАС. , Ли М.В. , Dell С. , Морган Л. , Молина П.Л. , Робинсон B.V. , Минникс S.L. , Ольбрих ЧАС. , Северин Т. , Аренс П. , Lange Л. , Мориллас H.N. , Никто П.Г. , Заривала М.А. и Ноулз МИСТЕР. , Врожденный порок сердца и другие гетеротаксические дефекты в большой группе пациентов с первичной цилиарной дискинезией, Circulation 115 (2007), 2814–2821. |
[77] | Ким R.H. , D А. , Cutz Э. , Ноулз МИСТЕР. , Неллиган К.А. , Nykamp К. , Заривала М.А. и Dell S.D. , Роль молекулярно-генетического анализа в диагностике первичной цилиарной дискинезии, Ann Am Thorac Soc 11 (2014), 351–359. |
[78] | Ноулз МИСТЕР. , Дэниелс L.A. , Дэвис S.D. , Заривала М.А. и Ли М.В. , Первичная цилиарная дискинезия.Последние достижения в диагностике, генетике и характеристике клинических заболеваний, Am J Respir Crit Care Med 188 (2013), 913–922. |
[79] | Ноулз МИСТЕР. , Ли М.В. , Карсон J.L. , Дэвис S.D. , Dell S.D. , Ферколь T.W. , Оливье К. , Сагель S.D. , Розенфельд М. , Ожоги К.А. , Минникс S.L. , Армстронг M.C. , Лори А. , Hazucha M.J. , Ложи N.T. , Ольбрих ЧАС. , Беккер-Хек А. , Шмидтс М. , Вернер С., Омран ЧАС. и Заривала М.А. , Мутации ДНКh21 у пациентов с первичной цилиарной дискинезией с нормальной ультраструктурой ресничек, Thorax 67 (2011), 433–441. |
[80] | Ноулз МИСТЕР. , Ли М.В. , Островский L.E. , Хуанг Л. , Карсон J.L. , Hazucha M.J. , Инь W. , Берг J.S. , Дэвис S.D. , Dell S.D. , Ферколь T.W. , Розенфельд М. , Сагель S.D. , Милла C.E. , Оливье К. , Тернер E.H. , Льюис А.П. , Бамшад М.Дж. , Никерсон Д.А. , Shendure Дж. , Заривала М.А. и Консорциум по генетическим нарушениям мукоцилиарного клиренса, секвенирование экзома идентифицирует мутации в CCDC114 как причину первичной цилиарной дискинезии, Am J Hum Genet 92 (2013), 99-106. |
[81] | Ноулз МИСТЕР. , Островский L.E. , ложи N.T. , Херд T.W. , Ли М. , Хуанг B.Q. , Волк МЫ. , Карсон J.L. , Hazucha M.J. , Инь W. , Дэвис S.D. , Dell S.D. , Ферколь Т., Сагель S.D. , Оливье К. , Янке С. , Ольбрих ЧАС. , Вернер С. , Raidt Дж. , Wallmeier Дж. , Pennekamp П. , Догерти G.W. , Hjeij Р. , Ну и дела H.Y. , Отто E.A. , Алебриттер Дж. , Чаки М. , Диас К.А. , Браун Д.А. , Порат J.D. , Schueler М. , Бактай ГРАММ. , Griese М. , Тернер E.H. , Льюис А.П. , Бамшад M.J. , Никерсон Д.А. , Хильдебрандт Ф. , Shendure Дж. , Омран ЧАС. и Заривала М. , Мутации в SPAG1 вызывают первичную дискинезию ресничек, ассоциированную с дефектными внешними и внутренними плечами динеина, Am J Hum Genet 93 (2013), 711-720. |
[82] | Коббернагель ОН. , Buchvald F.F. , Хаарман НАПРИМЕР. , Casaulta С. , Коллинз S.A. , Hogg С. , Куехни C.E. , Лукас J.S. , Омран ЧАС. , Quittner А.Л. , Вернер С. и Nielsen КГ. , Протокол исследования, обоснование и набор в европейское многоцентровое рандомизированное контролируемое исследование для определения эффективности и безопасности поддерживающей терапии азитромицином в течение 6 месяцев при первичной цилиарной дискинезии, BMC Pulm Med 16 (2016), 104. |
[83] | Котт Э. , Duquesnoy П. , Копин Б. , Legendre М. , Дасто-Ле Моаль Ф. , Монтантин Г. , Джинсон Л. , Тамалет А. , Папон Дж. Ф. , Сиффрой Дж. П. , Ривз Н. , Митчелл В. , де Блик Дж. , Косте А. , Клемент А. , Эскалье Д. , Туре А. , Эскудье Э. и Амселем С. , Мутации с потерей функции в LRRC6, гене, необходимом для правильной сборки аксонемы внутреннего и внешнего плеч динеина, вызывают первичную цилиарную дискинезию, Am J Hum Genet 91 (2012), 958–964. |
[84] | Куис П. , Папатеодору С.И. и Yiallouros П.К. , Диагностическая точность назального оксида азота для установления диагноза первичной цилиарной дискинезии: метаанализ, BMC Pulm Med 15 (2015), 153. |
[85] | Куехни C.E. , Фришер Т. , Стрипполи М.П. , Маурер Э. , куст А. , Nielsen КГ. , Escribano А. , Лукас J.S. , Yiallouros П. , Омран ЧАС., Эбер Э. , О’Каллаган С. , Снайдерс Д. , Барбато А. и Задача E.R.S. , Сила первичной цилиарной дискинезии у детей, Факторы, влияющие на возраст при диагностике первичной цилиарной дискинезии у европейских детей, Eur Respir J 36 (2010), 1248–1258. |
[86] | Ларкинс C.E. , Длинный А.Б. и Каспары Т. , Дефектная экспрессия Nodal и Cerl2 в мутантном узле Arl13b (hnn) лежит в основе его гетеротаксии, Dev Biol 367 (2012), 15-24. |
[87] | Ли М.В. , Ферколь T.W. , Дэвис S.D. , Ли H.S. , Розенфельд М. , Dell S.D. , Сагель S.D. , Милла С. , Оливье К. , Салливан К. , Заривала М.А. , Питтман J.E. , Шапиро А.Дж. , Карсон J.L. , Кришер Дж. , Hazucha M.J. и Ноулз МИСТЕР. , Клинические особенности и связанная с ними вероятность первичной цилиарной дискинезии у детей и подростков, Ann Am Thorac Soc 13 (2016), 1305–1313. |
[88] | Ли М.W. , Hazucha M.J. , Чавла К.К. , Бейкер Б.Р. , Шапиро А.Дж. , коричневый D.E. , Лаванж Л.М. , Horton Б.Дж. , Какиш Б. , Карсон J.L. , Дэвис S.D. , Dell S.D. , Ферколь T.W. , Аткинсон J.J. , Оливье К. , Сагель S.D. , Розенфельд М. , Милла С. , Ли H.S. , Кришер Дж. , Заривала М.А. и Ноулз МИСТЕР. , Стандартизация назального измерения оксида азота как теста на первичную цилиарную дискинезию, Ann Am Thorac Soc 10 (2013), 574–581. |
[89] | Ложи Н.Т. , Антоний Д. , Maver А. , Дирдорф М.А. , Gulec E.Y. , Гездиричи А. , Nothe-Menchen Т. , Hoben Я. , Jelten Л. , откровенный Д. , Вернер С. , Теббе Дж. , Ву К. , Гольдмунтц Э. , Кутурило ГРАММ. , Крок Б. , Риттер А. , Hjeij Р. , Бейки Z. , Pennekamp П. , Дворничак Б. , Бруннер ЧАС. , Петерлин Б. , Танидир С. , Ольбрих ЧАС. , Омран ЧАС. и Шмидтс М. , Рецессивные мутации потери функции DNAH9 вызывают дефекты латеральности и легкие респираторные дефекты биения ресничек, Am J Hum Genet 103 (2018), 995–1008. |
[90] | Ложи N.T. , Ольбрих ЧАС. , Беккер-Хек А. , Хаффнер К. , Heer А. , Рейнхард С. , Шмидтс М. , Kispert А. , Заривала М.А. , Ли М.В. , Ноулз МИСТЕР. , Zentgraf ЧАС. , Seithe ЧАС. , Нюрнберг ГРАММ. , Нюрнберг П. , Рейнхардт Р. и Омран ЧАС. , Делеции и точечные мутации LRRC50 вызывают первичную дискинезию ресничек из-за дефектов плеча динеина, Am J Hum Genet 85 (2009), 883–889. |
[91] | Ложи Н.Т. , Ольбрих ЧАС. , Фенске Л. , Муссаффи ЧАС. , Хорват Дж. , Fliegauf М. , Kuhl ЧАС. , Бактай ГРАММ. , Петерффи Э. , Чодхари Р. , Чанг ЭМ. , Рутман А. , О’Каллаган С. , Блау ЧАС. , Тиславич Л. , Voelkel К. , Витт М. , Зиткевич Э. , Neesen Дж. , Рейнхардт Р. , Mitchison H.M. и Омран ЧАС. , Мутации DNAI2 вызывают первичную цилиарную дискинезию с дефектами внешнего плеча динеина, Am J Hum Genet 83 (2008), 547–558. |
[92] | Мали ГРАММ.Р. , Йяти П.Л. , Mizuno С. , Додд ДЕЛАТЬ. , Теннант П.А. , Кейгрен М.А. , Зур Лаге П. , Шумарк А. , Гарсия-Муньос А. , Шимада А. , Такеда Х. , Эдлих Ф. , Такахаши С. , фон Крейгсхайм А. , Джарман А.П. и Милл П. , ZMYND10 функционирует в реле-шапероне во время сборки аксонемального динеина, Elife 7 (2018). |
[93] | Маршалл C.R. , Шерер С.В. , Заривала М.А. , Лау Л. , Патон Т.А. , Stockley Т. , Работа Р.К. , Луч П.Н. , Ноулз МИСТЕР. , Консорциум Forge Canada, зал Д.А. , Dell S.D. и Ким R.H. , Секвенирование всего экзома и анализ целевого числа копий при первичной цилиарной дискинезии, G3 (Bethesda) 5 (2015), 1775–1781. |
[94] | Мартин J.K. и Nielsen КГ. , Переносное носовое измерение оксида азота приливным дыханием — многообещающий инструмент для целевого выявления случаев для диагностики первичной цилиарной дискинезии, PLoS One 8 (2013), e57262. |
[95] | Мартин J.K. , Петерсен Н. , Сковгаард L.T. и Nielsen КГ. , Функция легких у пациентов с первичной цилиарной дискинезией: кросс-секционное и трехдесятилетнее продольное исследование, Am J Respir Crit Care Med 181 (2010), 1262–1268. |
[96] | Matwijiw Я. , Тливерис J.A. и Файман С. , Аплазия носовых ресничек с обратным положением, азооспермия и нормальные жгутики сперматозоидов: уникальный вариант синдрома неподвижных ресничек, J Urol 137 (1987), 522–524. |
[97] | Менер Д.Дж. , Линь С.Ю. , Ишман S.L. и Босс Э.Ф. , Лечение и исходы хронического риносинусита у детей с первичной цилиарной дискинезией: где доказательства? Качественный систематический обзор, Int Forum Allergy Rhinol 3 (2013), 986–991. |
[98] | Merveille A.C. , Дэвис E.E. , Беккер-Хек А. , Legendre М. , Амирав Я. , Батай ГРАММ., Бельмонт Дж. , Бейдон Н. , Биллен Ф. , Климент А. , Clercx С. , Косте А. , Кросби Р. , де Блик Дж. , Делёз С. , Дукесной П. , Эскалье Д. , Эскудье Э. , Флигауф М. , Хорват Дж. , Хилл К. , Йориссен М. , Просто Дж. , Кишперт А. , Латроп М. , Loges N.T. , Мартин Дж. К. , Момодзава Ю. , Монтантин Г. , Нильсен К.Г. , Ольбрих Х. , Папон Дж. Ф. , Райе И. , Роджер Г. , Шмидтс М. , Тенрейро Х. , Towbin J.A. , Зеленика Д. , Зентграф Х. , Жорж М. , Леквар А.С. , Кацанис Н. , Омран Х. и Амселем С. , CCDC39 необходим для сборки внутренних плеч динеина и регуляторного комплекса динеина и для нормальной подвижности ресничек у людей и собак, Nat Genet 43 (2011), 72–78. |
[99] | Мирра В. , Вернер С. и Санта Мария Ф. , Первичная цилиарная дискинезия: обновленная информация о клинических аспектах, генетике, диагностике и будущих стратегиях лечения, Front Pediatr 5 (2017), 135. |
[100] | Mitchison ЧАС.М. , Шмидтс М. , Ложи N.T. , Freshour Дж. , Dritsoula А. , Hirst Р.А. , О’Каллаган С. , Блау ЧАС. , Аль-Даббаг М. , Ольбрих Х. , Билс П.Л. , Яги Т. , Муссаффи Х. , Чунг Э. , Омран Х. и Митчелл Д. , Мутации в факторе сборки аксонемного динеина DNAAF3 вызывают первичную цилиарную дискинезию, Nat Genet 44 (2012), 381–389, S381–382. |
[101] | Мур А. , Escudier Э. , Роджер ГРАММ. , Тамалет А. , Pelosse Б., Марлин С. , Климент А. , Геремек М. , Delaisi Б. , Бриду ЯВЛЯЮСЬ. , Косте А. , Витт М. , Дурье Б. , Амселем С. , RPGR мутирует у пациентов со сложным X-сцепленным фенотипом, сочетающим первичную цилиарную дискинезию и пигментный ретинит, J Med Genet 43 (2006), 326–333. |
[102] | Мур Д.Дж. , Onoufriadis А. , Shoemark А. , Симпсон М.А. , Зур Лаге ЧИСЛО ПИ. , de Castro S.C. , Бартолони Л. , Галлоне Г. , Петриди С., Woollard W.J. , Энтони Д. , Шмидтс М. , Дидонна Т. , Макританасис П. , Бевиллард Дж. , Монган Н.П. , Дьяков Дж. , Пальс Г. , Лукас Дж. , Мартин Дж. К. , Нильсен К.Г. , Сантони Ф. , Гиппони М. , Хогг К. , Антонаракис С.Э. , Эмес Р.Д. , Чунг Э. , Грин Н.Д. , Блоуин Дж. Л. , Джарман А.П. и Митчисон Х. , Мутации в ZMYND10, гене, необходимом для правильной сборки аксонем внутреннего и внешнего плеч динеина у людей и мух, вызывают первичную цилиарную дискинезию, Am J Hum Genet 93 (2013), 346–356. |
[103] | Mullowney Т. , Мэнсон Д. , Ким Р. , Стивенс Д. , Шах В. и Dell С. , Первичная цилиарная дискинезия и респираторный дистресс у новорожденных, Педиатрия, 134 (2014), 1160–1166. |
[104] | Мюррей М.П. , Гован J.R. , Доэрти C.J. , Симпсон А.Дж. , Уилкинсон Т.С. , Чалмерс J.D. , Озеленение А.П. , Haslett С. и холм В. , Рандомизированное контролируемое исследование распыленного гентамицина при бронхоэктазах без кистозного фиброза, Am J Respir Crit Care Med 183 (2011), 491–499. |
[105] | Начуры М.В. , Молекулярные машины, передающие сигнальные рецепторы внутрь и из ресничек, Curr Opin Cell Biol 51 (2018), 124–131. |
[106] | Нонака С. , Танака Ю. , Окада Ю. , Такеда С. , Харада А. , Канаи Ю. , Кидо М. и Хирокава Н. , Рандомизация лево-правой асимметрии из-за потери узловых ресничек, генерирующих левый поток экстраэмбриональной жидкости у мышей, лишенных моторного белка KIF3B, Cell 95 (1998), 829-837. |
[107] | Никто П.Г. , Ли М.В. , Саннути А. , Минникс S.L. , Карсон J.L. , Hazucha М. , Заривала М.А. и Ноулз МИСТЕР. , Первичная цилиарная дискинезия: диагностические и фенотипические особенности, Am J Respir Crit Care Med 169 (2004), 459–467. |
[108] | О’Каллаган С. , Рутман А. , Уильямс Г. и Hirst Р.А. , Внутренние дефекты плеча динеина, вызывающие первичную цилиарную дискинезию: требуется повторное тестирование, Eur Respir J 38 (2011), 603–607. |
[109] | О’Доннелл A.E. , Баркер А.Ф. , Иловит J.S. и Фик R.B. , Лечение идиопатических бронхоэктазов с помощью аэрозольной рекомбинантной человеческой ДНКазы I. rhDNase Study Group, Chest 113 (1998), 1329–1334. |
[110] | Ольбрих ЧАС. , Хаффнер К. , Kispert А. , Volkel А. , Volz А. , Сасмаз ГРАММ. , Рейнхардт Р. , Хенниг С. , Lehrach ЧАС. , Конецко Н. , Заривала М., Никто П.Г. , Ноулз М. , Mitchison H.M. , Микс М. , Чанг ЭМ. , Хильдебрандт Ф. , Судбрак Р. и Омран ЧАС. , Мутации в DNAH5 вызывают первичную цилиарную дискинезию и рандомизацию лево-правой асимметрии, Nat Genet 30 (2002), 143–144. |
[111] | Ольбрих ЧАС. , Шмидтс М. , Вернер С. , Onoufriadis А. , Ложи N.T. , Raidt Дж. , Банки Н.Ф. , Shoemark А. , Burgoyne Т. , Аль Турки С. , Hurles M.E. , Колер Г., Шредер Дж. , Нюрнберг Г. , Нюрнберг П. , Чунг Э. , Рейнхардт Р. , Мартин Дж. К. , Нильсен К.Г. , Митчисон Х. и Омран Х. , Рецессивные мутации HYDIN вызывают первичную цилиарную дискинезию без рандомизации лево-правой асимметрии тела, Am J Hum Genet 91 (2012), 672–684. |
[112] | Olcese С. , Патель М.П. , Shoemark А. , Кивилуото С. , Legendre М. , Уильямс H.J. , Vaughan C.K. , Hayward Дж. , Гольденберг А., Emes R.D. , Munye М.М. , Красильщик Л. , Кэхилл Т. , Bevillard Дж. , Гериг С. , Гиппони М. , Chantot С. , Duquesnoy П. , Томас Л. , Джинсон Л. , Копин Б. , Тамалет А. , Товен-Робине С. , Папон J.F. , Гарин А. , Штырь Я. , Вера ГРАММ. , Аврора П. , Фассад МИСТЕР. , Дженкинс Л. , Бустред С. , Cullup Т. , Диксон М. , Onoufriadis А. , куст А. , Чанг ЭМ. , Антонаракис S.E. , Лёбингер МИСТЕР. , Уилсон Р. , Арменгот М. , Escudier Э. , Hogg С. , Группа U.K.R. , Амселем С. , солнце Z. , Бартолони Л. , Блуэн J.L. , Mitchison H.M. , Х-связанная первичная цилиарная дискинезия из-за мутаций в цитоплазматическом факторе сборки аксонемного динеина PIh2D3, Nat Commun 8 (2017), 14279. |
[113] | Олин J.T. , Ожоги К. , Карсон J.L. , Metjian ЧАС. , Аткинсон J.J. , Дэвис S.D. , Dell S.D. , Ферколь T.W. , Милла C.E. , Оливье К. , Розенфельд М. , Бейкер Б. , Ли М.В. , Ноулз М.Р. , Сагель S.D. и Консорциум по генетическим заболеваниям мукоцилиарного клиренса, Диагностический результат биопсии соскоба из носа при первичной цилиарной дискинезии: многоцентровый опыт, Pediatr Pulmonol 46 (2011), 483–488. |
[114] | Омран ЧАС. , Хаффнер К. , Volkel А. , Kuehr Дж. , Кетельсен ВВЕРХ. , Росс ЭМ-М-М. , Конецко Н. , Винкер Т. , Брэндис М. и Хильдебрандт Ф. , Картирование гомозиготности локуса гена первичной цилиарной дискинезии на хромосоме 5p и идентификация тяжелой цепи динеина DNAH5 в качестве гена-кандидата, Am J Respir Cell Mol Biol 23 (2000), 696–702. |
[115] | Омран ЧАС. , Кобаяши Д. , Ольбрих ЧАС. , Цукахара Т. , Ложи N.T. , Хагивара ЧАС. , Чжан В. , Леблон ГРАММ. , О’Тул Э. , Хара С. , Mizuno ЧАС. , Кавано ЧАС. , Fliegauf М. , Яги Т. , Кошида С. , Мияваки А. , Zentgraf ЧАС. , Seithe ЧАС. , Рейнхардт Р. , Ватанабэ Ю. , Камия Р. , Митчелл D.R. и Такеда ЧАС. , Ktu / PF13 необходим для цитоплазматической предварительной сборки аксонемных динеинов, Nature 456 (2008), 611-616. |
[116] | Onoufriadis А. , Пафф Т. , Антоний Д. , Shoemark А. , Миха Д. , Куйт Б. , Шмидтс М. , Петриди С. , Dankert-Roelse J.E. , Хаарман НАПРИМЕР. , Дэниелс J.M. , Emes R.D. , Уилсон Р. , Hogg С. , Скамблер П.Дж. , Чанг ЭМ. , Uk10K , Приятели ГРАММ. и Mitchison H.M. , Мутации сайтов сплайсинга в гене стыковочного комплекса внешнего динеинового плеча аксонемы CCDC114 вызывают первичную цилиарную дискинезию, Am J Hum Genet 92 (2013), 88–98. |
[117] | Onoufriadis А. , Shoemark А. , Шмидтс М. , Патель М. , Хименес ГРАММ. , Лю ЧАС. , Томас Б. , Диксон М. , Hirst Р.А. , Рутман А. , Burgoyne Т. , Уильямс С. , Скалли Дж. , Болард Ф. , Лафит J.J. , Beales П.Л. , Hogg С. , Ян П. , Чанг ЭМ. , Emes R.D. , О’Каллаган С. , Uk10K , Бувань П. и Mitchison H.M. , Панель целевых генов NGS идентифицирует мутации в RSPh2, вызывающие первичную цилиарную дискинезию и общий механизм агенеза центральной пары ресничек из-за дефектов радиальных спиц, Hum Mol Genet 23 (2014), 3362–3374. |
[118] | Пафф Т. , Дэниелс J.M. , Weersink E.J. , Lutter Р. , Вонк Нордеграф А. и Хаарман Э.Г. , Рандомизированное контролируемое исследование влияния ингаляционного гипертонического раствора на качество жизни при первичной цилиарной дискинезии, Eur Respir J 49 (2017). |
[119] | Пафф Т. , Ложи N.T. , Апреа Я. , Ву К. , Бейки Z. , Хаарман НАПРИМЕР. , Дэниелс J.M.A. , Sistermans Э.А. , Богунович Н. , Догерти G.W. , Hoben Я. , Гросс-Оннебринк Дж. , Иметь значение А. , Ольбрих ЧАС. , Вернер С. , Приятели ГРАММ. , Шмидтс М. , Омран ЧАС. , Миха Д. , Мутации в PIh2D3 вызывают Х-сцепленную первичную дискинезию ресничек с дефектами внешних и внутренних динеиновых плеч, Am J Hum Genet 100 (2017), 160–168. |
[120] | Приятель М. , Морган М. , Фелпс S.E. , Икра С.М. , Парри-Моррис С. , Дауны J.A. , Polier С. , Жемчужина Л.ЧАС. и Продромоу С. , Структурная основа зависимого от фосфорилирования рекрутирования Tel2 на Hsp90 с помощью Pih2, Structure 22 (2014), 805–818. |
[121] | Паницци J.R. , Беккер-Хек А. , Castleman V.H. , Аль-Мутаири Д.А. , Лю Ю. , Ложи N.T. , Патхак Н. , Остин-Цзе С. , Шеридан Э. , Шмидтс М. , Ольбрих ЧАС. , Вернер С. , Хаффнер К. , Хеллман Н. , Чодхари Р. , Гупта А. , Крамер-Цукер А. , Olale Ф. , Бурдин Р.Д. , Schier А.Ф. , О’Каллаган С. , Чанг ЭМ. , Рейнхардт Р. , Mitchison H.M. , король С.М. , Омран ЧАС. и Драммонд Я. , Мутации CCDC103 вызывают первичную дискинезию ресничек за счет нарушения сборки плеч цилиарного динеина, Nat Genet 44 (2012), 714-719. |
[122] | Пазур G.J. и Бладгуд Р.А. , Нацеливание белков на мембрану ресничек, Curr Top Dev Biol 85 (2008), 115–149. |
[123] | Pennarun ГРАММ., Escudier Э. , Чапелин С. , Бриду ЯВЛЯЮСЬ. , Cacheux В. , Роджер ГРАММ. , Климент А. , Goossens М. , Амселем С. и Дурье Б. , Мутации потери функции в человеческом гене, связанном с Chlamydomonas reinhardtii dynein IC78, приводят к первичной цилиарной дискинезии, Am J Hum Genet 65 (1999), 1508–1519. |
[124] | Пиффери М. , куст А. , Карамель Д. , Ди Чикко М. , Зангани М. , Чинеллато И. , Маккиа П. и Бонер А.Л. , Агенезия паральных пазух и оксид азота в носу при первичной цилиарной дискинезии, Eur Respir J 37 (2011), 566–571. |
[125] | Пиффери М. , Michelucci А. , Кониди МНЕ. , Кангиотти ЯВЛЯЮСЬ. , Сими П. , Macchia П. и Boner А.Л. , Новые мутации DNAh21 при первичной дискинезии ресничек с нормальной ультраструктурой аксонемы, Eur Respir J 35 (2010), 1413–1416. |
[126] | Пиперно ГРАММ.и Мид К. , Транспорт нового комплекса в цитоплазматическом матриксе жгутиков Chlamydomonas, Proc Natl Acad Sci U S. A 94 (1997), 4457–4462. |
[127] | Raidt Дж. , Вернер С. , Menchen Т. , Догерти G.W. , Ольбрих ЧАС. , Ложи N.T. , Шмитц Р. , Pennekamp П. и Омран ЧАС. , Функция ресничек и состав моторных белков фаллопиевых труб человека, Hum Reprod 30 (2015), 2871–2880. |
[128] | Ричард С., Незелоф С. , Пфистер А. , де Блик Дж. , Шейнманн П. и Пауп Ж. , Врожденная аплазия ресничек у двух братьев и сестер. Примитивное нарушение регуляции цилиогенеза? Патол Res Pract 185 (1989), 181–183. |
[129] | Россман СМ. , Форрест J.B. , Ли R.M. и Новый дом M.T. , Синдром дискинетических ресничек. Подвижность ресничек при синдроме неподвижных ресничек, Chest 78 (1980), 580–582. |
[130] | Rutland Дж.и Коул П. , Цилиарная дискинезия, Lancet 316 (1980), 859. |
[131] | Шах А. , Shoemark А. , MacNeill С.Дж. , Бхалудин Б. , Роджерс А. , Билтон Д. , Hansell D.M. , Уилсон Р. и Лёбингер МИСТЕР. , Продольное исследование, характеризующее большую популяцию взрослых с первичной цилиарной дискинезией, Eur Respir J 48 (2016), 441–450. |
[132] | Шах В КАЧЕСТВЕ. , Бен-Шахар Ю., Moninger К. , Kline J.N. и валлийский M.J. , Подвижные реснички эпителия дыхательных путей человека являются хемосенсорными, Science 325 (2009), 1131–1134. |
[133] | Шапиро А.Дж. , Дэвис S.D. , Ферколь Т. , Dell S.D. , Розенфельд М. , Оливье К. , Сагель S.D. , Милла С. , Заривала М.А. , Волк W. , Карсон J.L. , Hazucha M.J. , Ожоги К. , Робинсон Б. , Ноулз МИСТЕР. и Ли М.В. и Консорциум генетических нарушений мукоцилиарного клиренса, Латеральные дефекты, отличные от situs inversus totalis, при первичной цилиарной дискинезии: понимание situs ambiguus и гетеротаксии, Chest 146 (2014), 1176–1186. |
[134] | Шапиро А.Дж. , Дэвис S.D. , Полинени Д. , Manion М. , Розенфельд М. , Dell S.D. , Chilvers М.А. , Ферколь T.W. , Заривала М.А. , Сагель S.D. , Джозефсон М. , Морган Л. , Йылмаз О. , Оливье К. , Милла С. , Питтман J.E. , Дэниелс M.L.A. , Джонс M.H. , Джанахи Я. , Посуда С.М. , Даниэль С.Дж. , Купер М.Л. , Ноги Л.М. , Антон Б. , Eastvold Т. , Ehrne Л. , Guadagno Э. , Ноулз М.Р. , Ли М.В. , Лавернь В. и Ассамблея Американского торакального общества по педиатрии, Диагностика первичной дискинезии дилиарной железы. Официальное руководство по клинической практике Американского торакального общества, Am J Respir Crit Care Med 197 (2018), e24 – e39. |
[135] | Шапиро А.Дж. , Джозефсон М. , Розенфельд М. , Йылмаз О. , Дэвис S.D. , Полинени Д. , Guadagno Э. , Ли М.В. , Лавернь В. , Точность назального измерения оксида азота как диагностического теста первичной цилиарной дискинезии.Систематический обзор и метаанализ, Ann Am Thorac Soc 14 (2017), 1184–1196. |
[136] | Шапиро А.Дж. , Weck К.Э. , Чао К.С. , Розенфельд М. , Нигрен А.О. , Ноулз МИСТЕР. , Ли М.В. и Заривала М.А. , Синдром Кри-дю-чат и первичная цилиарная дискинезия: общая генетическая причина на хромосоме 5p, J Pediatr 165 (2014), 858–861. |
[137] | Шапиро А.Дж., Заривала М.А. , Ферколь Т. , Дэвис S.D. , Сагель S.D. , Dell S.D. , Розенфельд М. , Оливье К. , Милла С. , Даниэль С.Дж. , Kimple А.Дж. , Manion М. , Ноулз МИСТЕР. , Ли М.В. и Консорциум по генетическим заболеваниям мукоцилиарного клиренса, Диагностика, мониторинг и лечение первичной цилиарной дискинезии: согласованные рекомендации Фонда PCD, основанные на обзоре современного состояния, Pediatr Pulmonol 51 (2016), 115–132. |
[138] | Shoemark А., Burgoyne Т. , Кван Р. , Диксон М. , Патель М.П. , Роджерс СРЕДНИЙ. , Onoufriadis А. , Скалли Дж. , Даудвохра Ф. , Cullup Т. , Лёбингер МИСТЕР. , Уилсон Р. , Чанг E.M.K. , куст А. , Mitchison H.M. и Hogg С. , Первичная цилиарная дискинезия с нормальной ультраструктурой: трехмерная томография выявляет отсутствие ДНКh21, Eur Respir J 51 (2018). |
[139] | Shoemark А. , Диксон М. , Коррин Б. и Дьюар А., Двадцатилетний обзор количественной просвечивающей электронной микроскопии для диагностики первичной цилиарной дискинезии, J Clin Pathol 65 (2012), 267–271. |
[140] | Симет С.М. , Сиссон J.H. , Павлик J.A. , Devasure J.M. , Boyer С. , Лю ИКС. , Кавасаки С. , Острый J.G. , Реннард С.И. и Wyatt Т.А. , Длительное воздействие сигаретного дыма на мышиной модели функции мерцательных эпителиальных клеток, Am J Respir Cell Mol Biol 43 (2010), 635–640. |
[141] | Сани М.А. , Первичная цилиарная дискинезия, Lancet 2 (1981), 476. |
[142] | Smit H.J. , Schreurs А.Дж. , Ван ден Бош J.M. и Вестерманн К.Дж. , Полезна ли резекция бронхоэктазов у пациентов с первичной цилиарной дискинезией? Сундук 109 (1996), 1541–1544. |
[143] | Sommer J.U., Шафер К. , Омран ЧАС. , Ольбрих ЧАС. , Wallmeier Дж. , Блюм А. , Хорманн К. и Застрявший Б.А. , ЛОР-проявления у пациентов с первичной цилиарной дискинезией: распространенность и значение сопутствующих оториноларингологических заболеваний, Eur Arch Otorhinolaryngol 268 (2011), 383–388. |
[144] | Сорокин С.П. , Реконструкции образования центриолей и цилиогенеза в легких млекопитающих, J Cell Sci 3 (1968), 207–230. |
[145] | Таркар А., Ложи N.T. , Slagle C.E. , Фрэнсис Р. , Догерти G.W. , Тамайо J.V. , Потряс Б. , Кантино М. , Шварц Д. , Янке С. , Ольбрих ЧАС. , Вернер С. , Raidt Дж. , Pennekamp П. , Абухамед М. , Hjeij Р. , Колер ГРАММ. , Griese М. , Ли Ю. , Лемке К. , Клена Н. , Лю ИКС. , Габриэль ГРАММ. , Тобита К. , Ясперс М. , Морган L.C. , Шапиро А.Дж. , Letteboer С.Дж. , Ман Д.А. , Карсон J.L. , Ли М.В. , Волк МЫ. , Чен С. , Лукас J.S. , Onoufriadis А., Plagnol В. , Шмидтс М. , Boldt К. , Uk10K, Роепман Р. , Заривала М.А. , Lo C.W. , Митчисон Х. , Ноулз М. , Бурдин Р.Д. , Loturco J.J. и Омран Х. , DYX1C1 необходим для сборки аксонемального динеина и подвижности ресничек, Nat Genet 45 (2013), 995–1003. |
[146] | Theegarten Д. и Эбсен М. , Ультраструктурная патология первичной цилиарной дискинезии: Сообщите о 125 случаях в Германии, Diagn Pathol 6 (2011), 115. |
[147] | Тобин J.L. и Beales П.Л. , Неподвижные цилиопатии, Genet Med 11 (2009), 386–402. |
[148] | Wallmeier Дж. , Аль-Мутаири Д.А. , Чен C.T. , Ложи N.T. , Pennekamp П. , Menchen Т. , Ма Л. , Шамсельдин ОН. , Ольбрих ЧАС. , Догерти G.W. , Вернер С. , Альсабах B.H. , Колер ГРАММ. , Ясперс М. , Благо М. , Griese М. , Шмитт-Гроэ С., Циммерманн Т. , Кёрнер-Реттберг С. , Горак Э. , Кинтнер С. , Алькурая Ф.С. и Омран ЧАС. , Мутации в CCNO приводят к врожденному нарушению мукоцилиарного клиренса с уменьшением генерации множественных подвижных ресничек, Nat Genet 46 (2014), 646–651. |
[149] | Wallmeier Дж. , Ширатори ЧАС. , Догерти G.W. , Эдельбуш С. , Hjeij Р. , Ложи N.T. , Menchen Т. , Ольбрих ЧАС. , Pennekamp П. , Raidt Дж. , Вернер С., Минегиси К. , Шинохара К. , Асаи Ю. , Такаока К. , Ли С. , Griese М. , Мемари Ю. , Дурбин Р. , Кольб-Кокочинский А. , Зауэр С. , Wallingford J.B. , Хамада ЧАС. и Омран ЧАС. , Дефицит TTC25 приводит к дефектам аппарата стыковки внешнего плеча динеина и первичной цилиарной дискинезии с рандомизацией лево-правой асимметрии тела, Am J Hum Genet 99 (2016), 460–469. |
[150] | Посуда С.М. , Айгюн М.Г. и Хильдебрандт Ф., Спектр клинических заболеваний, вызванных нарушениями первичных ресничек, Proc Am Thorac Soc 8 (2011), 444–450. |
[151] | Ватанабэ Д. , Сайджох Ю. , Нонака С. , Сасаки ГРАММ. , Икава Ю. , Ёкояма Т. и Хамада ЧАС. Лево-правый детерминант Inversin является компонентом моноцилий узла и др. 9 + 0 ресничек, Development 130 (2003), 1725-1734. |
[152] | Wijers С.Д. , Chmiel J.F. и Гастон Б.М. , Бактериальные инфекции у пациентов с первичной цилиарной дискинезией: сравнение с муковисцидозом, Chron Respir Dis 14 (2017), 392–406. |
[153] | Вонг С. , Джаярам Л. , Каралус Н. , Eaton Т. , Тонг С. , Хоккей ЧАС. , Milne Д. , Фергюссон W. , Туффери С. , Секстон П. , Этаж Л. и Эштон Т. , Азитромицин для профилактики обострений бронхоэктатической болезни без кистозного фиброза (EMBRACE): рандомизированное двойное слепое плацебо-контролируемое исследование, Lancet 380 (2012), 660–667. |
[154] | Ян П. , Динер D.R. , Ян С. , Коно Т. , Пазур G.J. , Диены J.M. , Усмешка Н.С. , король С.М. , распродажа W.S. , Камия Р. , Розенбаум J.L. и Витман Дж. Б. , Радиально-спицевые белки жгутиков Chlamydomonas, J Cell Sci 119 (2006), 1165–1174. |
[155] | Заривала М.А. , Ну и дела H.Y. , Курковяк М. , Аль-Мутаири Д.А. , Ли М.В. , Херд Т.W. , Hjeij Р. , Dell S.D. , Чаки М. , Догерти G.W. , Адан М. , Копье ПК. , Эстев-Радд Дж. , Ложи N.T. , Розенфельд М. , Диас К.А. , Ольбрих ЧАС. , Волк МЫ. , Шеридан Э. , Планка Т.Ф. , Алебриттер Дж. , Порат J.D. , Коль С. , Ловрич С. , Hwang Д.Ю. , Питтман J.E. , Ожоги К.А. , Ферколь T.W. , Сагель S.D. , Оливье К. , Морган L.C. , Вернер С. , Raidt Дж. , Pennekamp П. , солнце Z. , Чжоу W. , Айрик Р. , Натараджан С. , Аллен С.Дж. , Амирав Я., Wieczorek Д. , Ландвер К. , Nielsen К. , Schwerk Н. , Sertic Дж. , Колер ГРАММ. , Washburn Дж. , Леви С. , Поклонник С. , Кёрнер-Реттберг С. , Амселем С. , Уильямс D.S. , Митчелл Б.Дж. , Драммонд Я. , Отто E.A. , Омран ЧАС. , Ноулз МИСТЕР. и Хильдебрандт Ф. , ZMYND10 мутирует при первичной цилиарной дискинезии и взаимодействует с LRRC6, Am J Hum Genet 93 (2013), 336–345. |
[156] | Заривала М.А. , Ли М.W. , Чеппа Ф. , Кеннеди М.П. , Никто П.Г. , Карсон J.L. , Hazucha M.J. , Лори А. , Хорват Дж. , Ольбрих ЧАС. , Ложи N.T. , Бриду ЯВЛЯЮСЬ. , Pennarun ГРАММ. , Дурье Б. , Escudier Э. , Mitchison H.M. , Чодхари Р. , Чанг ЭМ. , Морган L.C. , де Ионг RU. , Ратленд Дж. , Прадал У. , Омран Х. , Амселем С. и Ноулз М. Мутации ДНКI1 при первичной цилиарной дискинезии: доказательство эффекта основателя при общей мутации, Am J Respir Crit Care Med 174 (2006), 858–866. |
[157] | Заривала М.А. , Омран ЧАС. и Ферколь T.W. , Возникающая генетика первичной цилиарной дискинезии, Proc Am Thorac Soc 8 (2011), 430–433. |
 А.
, Синдром человека, вызванный неподвижными ресничками, Science 193 (1976), 317–319.
А.
, Синдром человека, вызванный неподвижными ресничками, Science 193 (1976), 317–319.

 ,
Ложи
N.T.
,
Ольбрих
ЧАС.
,
Haeffner
К.
,
Fliegauf
М.
,
Хорват
Дж.
,
Рейнхардт
Р.,
Nielsen
КГ.
,
Мартин
J.K.
,
Бактай
ГРАММ.
,
Андерсон
К.В.
,
Гейслер
Р.
,
Niswander
Л.
,
Омран
ЧАС.
и
Бурдин
R.D.
Домен coiled-coil, содержащий белок CCDC40, важен для функции подвижных ресничек и формирования оси слева направо, Nat Genet 43 (2011), 79–84.
,
Ложи
N.T.
,
Ольбрих
ЧАС.
,
Haeffner
К.
,
Fliegauf
М.
,
Хорват
Дж.
,
Рейнхардт
Р.,
Nielsen
КГ.
,
Мартин
J.K.
,
Бактай
ГРАММ.
,
Андерсон
К.В.
,
Гейслер
Р.
,
Niswander
Л.
,
Омран
ЧАС.
и
Бурдин
R.D.
Домен coiled-coil, содержащий белок CCDC40, важен для функции подвижных ресничек и формирования оси слева направо, Nat Genet 43 (2011), 79–84. ,
де Блик
Дж.
,
Климент
А.
,
Escudier
Э.
и
Амселем
С.
, Определение спектра мутаций CCDC39 / CCDC40 и связанных фенотипов при первичной цилиарной дискинезии, J Med Genet 49 (2012), 410–416.
,
де Блик
Дж.
,
Климент
А.
,
Escudier
Э.
и
Амселем
С.
, Определение спектра мутаций CCDC39 / CCDC40 и связанных фенотипов при первичной цилиарной дискинезии, J Med Genet 49 (2012), 410–416.


 ,
Паркер
К.А.
,
Ибот-Гонсалес
П.
,
Emes
R.D.
,
Уилсон
С.В.
,
Уоллис
С.
,
Джонсон
C.A.
,
Эррера
Р.Дж.
,
Рутман
А.,
Диксон
М.
,
Shoemark
А.
,
куст
А.
,
Hogg
С.
,
Гардинер
R.M.
,
Рейш,
О.
,
Грин
N.D.
,
О’Каллаган
С.
,
Purton
С.
,
Чанг
ЭМ.
и
Mitchison
H.M.
, Мутации в генах белков RSPH9 и RSPh5A радиальной спицевой головы вызывают первичную дискинезию ресничек с аномалиями пары центральных микротрубочек, Am J Hum Genet 84 (2009), 197–209.
,
Паркер
К.А.
,
Ибот-Гонсалес
П.
,
Emes
R.D.
,
Уилсон
С.В.
,
Уоллис
С.
,
Джонсон
C.A.
,
Эррера
Р.Дж.
,
Рутман
А.,
Диксон
М.
,
Shoemark
А.
,
куст
А.
,
Hogg
С.
,
Гардинер
R.M.
,
Рейш,
О.
,
Грин
N.D.
,
О’Каллаган
С.
,
Purton
С.
,
Чанг
ЭМ.
и
Mitchison
H.M.
, Мутации в генах белков RSPH9 и RSPh5A радиальной спицевой головы вызывают первичную дискинезию ресничек с аномалиями пары центральных микротрубочек, Am J Hum Genet 84 (2009), 197–209.
 Ed (2018).
Ed (2018).
 W.
,
Ложи
N.T.
,
Клинкенбуш
J.A.
,
Ольбрих
ЧАС.
,
Pennekamp
П.
,
Menchen
Т.
,
Raidt
Дж.
,
Wallmeier
Дж.
,
Вернер
С.
,
Вестерманн
С.
,
Рукерт
С.
,
Мирра
В.
,
Hjeij
Р.
,
Мемари
Ю.
,
Дурбин
Р.
,
Кольб-Кокочинский
А.
,
Правин
К.,
Кашеф
М.А.
,
Кашеф
С.
,
Эхтедари
Ф.
,
Хаффнер
К.
,
Валмари
П.
,
Бактай
ГРАММ.
,
Авирам
М.
,
Бентур
Л.
,
Амирав
Я.
,
Дэвис
E.E.
,
Кацанис
Н.
,
Брюкнер
М.
,
Шапошников
А.
,
Пигино
ГРАММ.
,
Дворничак
Б.
и
Омран
ЧАС.
, Локализация DNAh21 в проксимальной области респираторных ресничек определяет отдельные комплексы внешних динеиновых плеч, Am J Respir Cell Mol Biol 55 (2016), 213–224.
W.
,
Ложи
N.T.
,
Клинкенбуш
J.A.
,
Ольбрих
ЧАС.
,
Pennekamp
П.
,
Menchen
Т.
,
Raidt
Дж.
,
Wallmeier
Дж.
,
Вернер
С.
,
Вестерманн
С.
,
Рукерт
С.
,
Мирра
В.
,
Hjeij
Р.
,
Мемари
Ю.
,
Дурбин
Р.
,
Кольб-Кокочинский
А.
,
Правин
К.,
Кашеф
М.А.
,
Кашеф
С.
,
Эхтедари
Ф.
,
Хаффнер
К.
,
Валмари
П.
,
Бактай
ГРАММ.
,
Авирам
М.
,
Бентур
Л.
,
Амирав
Я.
,
Дэвис
E.E.
,
Кацанис
Н.
,
Брюкнер
М.
,
Шапошников
А.
,
Пигино
ГРАММ.
,
Дворничак
Б.
и
Омран
ЧАС.
, Локализация DNAh21 в проксимальной области респираторных ресничек определяет отдельные комплексы внешних динеиновых плеч, Am J Respir Cell Mol Biol 55 (2016), 213–224. ,
ас-Сархани
А.
и
аль-Эсса
A.R.
, Отологические проявления первичной цилиарной дискинезии, Clin Otolaryngol Allied Sci 22 (1997), 266–270.
,
ас-Сархани
А.
и
аль-Эсса
A.R.
, Отологические проявления первичной цилиарной дискинезии, Clin Otolaryngol Allied Sci 22 (1997), 266–270.Частота биений ресничек трахеи у павианов: эффекты периферического гистамина и капсаицина
Д. Б. Йейтс, Б. Р. Питт, Д. М. Спектор, Г. А. Каррон и Р. Э. Альберт, Координация мукоцилиарного транспорта в трахее человека и внутрилегочных дыхательных путей .J. Appl. Physiol. 51, , 1057–1064 (1981).
PubMed Google ученый
Д. Б. Йейтс, Т. Р. Геррити и К. С. Гаррард, Характеристики трахеобронхиального отложения и клиренса у человека . Анна. Ок. Hyg. 26 , 245–257 (1982).
PubMed Google ученый
А. Ваннер, Д. Маурер, В. М. Абрахам, З. Сепфалуси и М.Sielczak, Влияние химических медиаторов анафилаксии на функцию ресничек . J. Allergy Clin. Иммунол. 72 , 663–667 (1983).
PubMed Google ученый
Х. Бисгаард и М. Педерсен, Лейкотриены SRS-A снижают активность респираторных ресничек человека . Clin. Аллергия. 17, , 95–103 (1987).
PubMed Google ученый
Дж.В. Скади, Э. Т. Кумура и Дж. Ф. Рейнхард, Исследование действия лекарств на мерцательный эпителий млекопитающих . J. Pharmacol. Exper. Ther. 102 , 132–137 (1951).
Google ученый
К. С. Гаррад, Д. Муссатто и Р. В. Лоренко, Мукоцилиарный транспорт легких при бессимптомной астме: эффекты вдыхаемого гистамина . J. Lab. Clin. Med. 113 , 190–195 (1989).
PubMed Google ученый
D.Дж. Муссатто, К. С. Гаррард и Р. В. Лоренко, Влияние вдыхаемого гистамина на скорость слизи в трахее человека и мукоцилиарный клиренс бронхов . Являюсь. Преподобный Респир. Дис. 138 , 775–779 (1988).
PubMed Google ученый
N. Chand, Распределение и классификация гистаминовых рецепторов дыхательных путей: физиологическое значение гистамина H 2 — рецепторов . Adv.Pharm. Химиотерапия 17 , 103–131 (1980).
Google ученый
Х. М. Кольридж, Дж. С. Г. Кольридж, К. Гинзель, Д. Бейкер, Р. Банцетт и М. Моррисон, Стимуляция «раздражающих» рецепторов и афферентных С-волокон в легких простагландинами . Nature 264 , 451–453 (1976).
PubMed Google ученый
Л. Б. Вонг, И.Ф. Миллер и Д. Б. Йейтс, Стимуляция частоты биений ресничек трахеи капсаицином . J. Appl. Physiol. 68 , 2574–2580 (1990).
PubMed Google ученый
Р. Носаль, К. Драбикова, Ю. Печивова и Дж. Якубовский, Высвобождение гистамина в результате нерецепторного взаимодействия . Агенты и действия 30 , 74–76 (1990).
PubMed Google ученый
B.Фокс, Т. Б. Булл и А. Газ, Тучные клетки в альвеолярной стенке человека: электронно-микроскопическое исследование . J. Clin. Патол. 34 , 1333–1342, 1981.
PubMed Google ученый
Л. Б. Вонг, И. Ф. Миллер и Д. Б. Йейтс, Стимуляция частоты биений ресничек вегетативными агонистами: in vivo . J. Appl. Physiol. 65 , 971–981 (1988).
PubMed Google ученый
S.A. Sirr, P. J. Juenemann, H. Tom, R. J. Boudreau, R. P. Chandler и M. K. Loken, Влияние этанола на размер капель, эффективность доставки и характеристики клиренса аэрозоля DTPA технеция-99m . J. Nucl Med. 26, , 643–646 (1985).
PubMed Google ученый
Дж. Ратленд и П. Дж. Коул, Неинвазивный отбор проб носовых ресничек для измерения частоты биений и исследования ультраструктуры .Lancet , 13 сентября, , 564–565 (1980).
Google ученый
С. Янаура, Н. Имамура и М. Мисава, Эффекты стимуляторов β-адренорецепторов на реснитчатые клетки трахеи собак . Japan J. Pharmacol. 31, , 951–956 (1981).
Google ученый
Дж. Ягер, Т. Чен и М. Дульфано, Измерение частоты цилиарных сокращений респираторного эпителия человека .Сундук 73 , 627–633 (1978).
PubMed Google ученый
Л. Б. Вонг, И. Ф. Миллер и Д. Б. Йейтс, Регуляция частоты биений ресничек с помощью вегетативных механизмов: in vitro . J. Appl. Physiol. 65 , 1895–1901 (1988).
PubMed Google ученый
S. Lindberg и U. Mercke, Антагонисты вещества P и мукоцилиарная активность у кроликов .Arch Pharmacol Наунин-Шмидеберг. 329 , 376–381 (1985).
Google ученый
Л. Б. Вонг и Д. Б. Йейтс, Стимуляция частоты биений ресничек трахеи посредством локализованного разреза ткани . J. Appl. Physiol. 68 , 411–416 (1990).
PubMed Google ученый
В. Лоренц, В. Дитц, М. Эннис, Б. Стиннер и А. Денике, Гистамин в анестезии и хирургии: анализ причинной связи .В Справочнике по экспериментальной фармакологии : гистамин и антагонисты гистамина , vol. 97. (Ed. B. Uvnas) pp. 385–439, Springer-Verlag, Berlin 1991.
Google ученый
Э. Родригес-Булан и У. Дж. Нельсон, Морфогенез фенотипа поляризованных эпителиальных клеток . Science 245 , 718–725 (1989).
PubMed Google ученый
М.Липпман и Р. Э. Альберт, Влияние размера частиц на региональное осаждение вдыхаемых аэрозолей в дыхательных путях человека . Являюсь. Ind. Hyg. Доц. J. 30 , 257–275 (1969).
PubMed Google ученый
Дж. Хейдер, Альвеолярное отложение вдыхаемых частиц у людей . Являюсь. Ind. Hyg. Доц. J. 43 , 864–866 (1982).
PubMed Google ученый
М.Eljamal, L. B. Wong и D. B. Yeates, Периферийное отложение капсаицина в аэрозольной форме стимулирует частоту биений ресничек трахеи собак как через симпатические, так и через парасимпатические пути . Являюсь. Преподобный Респир. Дис. 141 , ( 4 ). A847 (1990 г.).
Google ученый
М. Нисам, А. Збинден, С. Чесроун, Д. Барнетт и В. Голд, Распределение и фармакологическое высвобождение гистамина в легких собаки in vivo .J. Appl. Physiol. 44 , 455–463 (1978).
PubMed Google ученый
Н. Чанд, Б. Н. Дхаван, Р. К. Сримал, Н. Х. Рахмани, Р. Шукла и Б. М. Алтура, Реакция трахеи, бронхов и легких полосок на гистамин и карбахол у макак-резусов . J. Appl. Physiol. 49 , 729–734 (1980).
PubMed Google ученый
Дж.А. Надель, М. Корн, С. Цви, Дж. Флеш и П. Граф, Местоположение и механизм сужения дыхательных путей после вдыхания аэрозоля гистамина и аэрозоля органических сульфатов . В Вдыхаемых частиц и паров II . Proc. Международного Сим. Брит. Ок. Hyg. Soc., Стр. 55–67, Pergamon Press, Oxford 1967.
Google ученый
А. Ваннер, Дж. М. Броднан, Дж. Перес, К. Г. Хенке и К. С. Ким, Вариабельность реакции дыхательных путей на аэрозоль гистамина у здоровых субъектов .Являюсь. Преподобный Респир. Дис. 131 , 3–7 (1985).
PubMed Google ученый
Н. К. Барнс, П. Пайпер и Дж. Костелло, Сравнительные эффекты вдыхаемого лейкотриена С 4 , лейкотриен D 4 , и гистамин у нормальных людей . Грудь 39 , 500–504 (1984).
PubMed Google ученый
р.В. Фуллер, К. М. С. Диксон и П. Дж. Барнс, Бронхоконстрикторный ответ на вдыхаемый капсаицин у людей . J. Appl. Physiol. 58, , 1080–1084 (1985).
PubMed Google ученый
Р. Носаль, К. Драбикова, Дж. Печивова и К. Ондриас, Мембранно-возмущающая активность препаратов, блокирующих бета-адренорецепторы, в изолированных тучных клетках крысы . Агенты и действия 27 , 36–38 (1989).
PubMed Google ученый
Дж.Мало, Р. Готер, И. Лемир, А. Картье, Х. Геццо и Р. Р. Мартин, Кинетика восстановления реакции дыхательных путей, вызванной вдыханием гистамина . Являюсь. Преподобный Респир. Дис. 132 , 848–852 (1985).
PubMed Google ученый
Реснички, полосатые мыши и ожирение | Реснички
Whitfield JF: Нейрональная первичная ресничка — внесинаптическое сигнальное устройство. Сотовый сигнал. 2004, 16: 763-767. 10.1016 / j.cellsig.2003.12.002.
CAS Статья Google ученый
Хильдебрандт Ф., Бенцинг Т., Катсанис Н .: Цилиопатии. N Engl J Med. 2011, 364: 1533-1543. 10.1056 / NEJMra1010172.
CAS Статья Google ученый
Билс П.Л., Фаруки И.С., О’Рахилли С. Генетика синдромов ожирения. 2009, Нью-Йорк: Oxford University Press
Google ученый
Давенпорт JR, Уоттс AJ, Roper VC, Croyle MJ, van Groen T, Wyss JM, Nagy TR, Kesterson RA, Yoder BK: Нарушение внутрилагеллярного транспорта у взрослых мышей приводит к ожирению и медленному развитию кистозной болезни почек. Curr Biol. 2007, 17: 1586-1594. 10.1016 / j.cub.2007.08.034.
CAS Статья Google ученый
Coleman DL, Eicher EM: Fat (жир) и tubby (tub): две аутосомно-рецессивные мутации, вызывающие синдромы ожирения у мышей.J Hered. 1990, 81: 424-427.
CAS Google ученый
Kleyn PW, Fan W, Kovats SG, Lee JJ, Pulido JC, Wu Y, Berkemeier LR, Misumi DJ, Holmgren L, Charlat O, Woolf EA, Tayber O, Brody T, Shu P, Hawkins F , Kennedy B, Baldini L, Ebeling C, Alperin GD, Deeds J, Lakey ND, Culpepper J, Chen H, Glücksmann-Kuis MA, Carlson GA, Duyk GM, Moore KJ: Идентификация и характеристика гена ожирения мышей tubby: a член семейства генов anovel.Клетка. 1996, 85 (2): 281-290. 10.1016 / S0092-8674 (00) 81104-6.
CAS Статья Google ученый
Нобен-Траут К., Наггерт Дж. К., Северный Массачусетс, Нишина П.М.: ген-кандидат для мутации мышей тубби. Природа. 1996, 380: 534-538. 10.1038 / 380534a0.
CAS Статья Google ученый
Ashrafi K, Chang FY, Watts JL, Fraser AG, Kamath RS, Ahringer J, Ruvkun G: Полногеномный РНКи-анализ генов регуляции жиров Caenorhabditis elegans.Природа. 2003, 421: 268-272. 10.1038 / природа01279.
CAS Статья Google ученый
Мак Х.Й., Нельсон Л.С., Бассон М., Джонсон С.Д., Рувкун Г.: Полигенный контроль накопления жира Caenorhabditis elegans. Нат Жене. 2006, 38: 363-368. 10.1038 / ng1739.
CAS Статья Google ученый
Sun X, Haley J, Bulgakovoleg OV, Cai X, McGinnis J, Li T: Табби необходим для доставки рецепторов, связанных с g-белком, в нейрональные цилии.Реснички. 2012, 1: 21-10.1186 / 2046-2530-1-21.
CAS Google ученый
Handel M, Schulz S, Stanarius A, Schreff M, Erdtmann-Vourliotis M, Schmidt H, Wolf G, Hollt V: Селективное нацеливание рецептора 3 соматостатина на реснички нейронов. Неврология. 1999, 89: 909-926. 10.1016 / S0306-4522 (98) 00354-6.
CAS Статья Google ученый
Berbari NF, Lewis JS, Bishop GA, Askwith CC, Mykytyn K: белки синдрома Барде-Бидла необходимы для локализации рецепторов, связанных с G-белком, в первичных ресничках.Proc Natl Acad Sci USA. 2008, 105: 4242-4246. 10.1073 / pnas.0711027105.
CAS Статья Google ученый
Kulaga HM, Leitch CC, Eichers ER, Badano JL, Lesemann A, Hoskins BE, Lupski JR, Beales PL, Reed RR, Katsanis N: Потеря белков BBS вызывает аносмию у людей и дефекты в структуре обонятельной цилиоструктуры и функции в мыши. Нат Жене. 2004, 36: 994-998. 10.1038 / ng1418.
CAS Статья Google ученый
Мукхопадхьяй С., Джексон П.К .: Белки семейства толстых. Genome Biol. 2011, 12: 225-10.1186 / gb-2011-12-6-225.
CAS Статья Google ученый
Santagata S, Boggon TJ, Baird CL, Gomez CA, Zhao J, Shan WS, Myszka DG, Shapiro L: передача сигналов G-белка через белки тубби. Наука. 2001, 292: 2041-2050. 10.1126 / science.1061233.
CAS Статья Google ученый
Mukhopadhyay S, Wen X, Chih B, Nelson CD, Lane WS, Scales SJ, Jackson PK: TULP3 связывает комплекс IFT-A и мембранные фосфоинозитиды, способствуя перемещению рецепторов, связанных с G-белком, в первичные реснички. Genes Dev. 2010, 24: 2180-2193. 10.1101 / gad.1966210.
CAS Статья Google ученый
Norman RX, Ko HW, Huang V, Eun CM, Abler LL, Zhang Z, Sun X, Eggenschwiler JT: Tubby-подобный белок 3 (TULP3) регулирует формирование паттерна в эмбрионе мыши посредством ингибирования передачи сигналов Hedgehog.Hum Mol Genet. 2009, 18: 1740-1754. 10.1093 / hmg / ddp113.
CAS Статья Google ученый
Шимада М., Тритос Н.А., Лоуэлл Б.Б., Флиер Дж. С., Маратос-Флиер Э. Мыши, у которых отсутствует гормон, концентрирующий меланин, гипофагичны и худощавы. Природа. 1998, 396: 670-674. 10.1038 / 25341.
CAS Статья Google ученый
Chen Y, Hu C, Hsu CK, Zhang Q, Bi C, Asnicar M, Hsiung HM, Fox N, Slieker LJ, Yang DD, Heiman ML, Shi Y: Целенаправленное нарушение выработки меланин-концентрирующего гормона. рецептор-1 приводит к гиперфагии и устойчивости к ожирению, вызванному диетой.Эндокринология. 2002, 143: 2469-2477. 10.1210 / en.143.7.2469.
CAS Статья Google ученый
Людвиг Д.С., Тритос Н.А., Мастайтис Дж. В., Кулкарни Р., Коккоту Э., Элмквист Дж., Лоуэлл Б., Флиер Дж. С., Маратос-Флиер Е. Сверхэкспрессия меланин-концентрирующего гормона у трансгенных мышей приводит к ожирению и инсулинорезистентности. J Clin Invest. 2001, 107: 379-386. 10.1172 / JCI10660.
CAS Статья Google ученый
Wang Z, Li V, Chan GC, Phan T, Nudelman AS, Xia Z, Storm DR: Взрослые мыши с дефицитом аденилилциклазы 3 типа страдают ожирением. PLoS One. 2009, 4: e6979-10.1371 / journal.pone.0006979.
Артикул Google ученый
Hawes BE, Kil E, Green B, O’Neill K, Fried S, Graziano MP: Рецептор меланин-концентрирующего гормона соединяется с множеством G-белков для активации различных внутриклеточных сигнальных путей. Эндокринология. 2000, 141: 4524-4532.10.1210 / en.141.12.4524.
CAS Статья Google ученый
Pissios P, Trombly DJ, Tzameli I., Maratos-Flier E: Рецептор 1 гормона, концентрирующего меланин, активирует киназу, регулируемую внеклеточными сигналами, и взаимодействует с G (s) -связанными путями. Эндокринология. 2003, 144: 3514-3523. 10.1210 / en.2002-0004.
CAS Статья Google ученый
Marley A, von Zastrow M: DISC1 регулирует первичные реснички, которые отображают специфические рецепторы дофамина.PLoS One. 2010, 5: e10902-10.1371 / journal.pone.0010902.
Артикул Google ученый
Goetz SC, Андерсон К.В.: Первичная ресничка: центр передачи сигналов во время развития позвоночных. Nat Rev Genet. 2010, 11: 331-344. 10.1038 / nrg2774.
CAS Статья Google ученый
Чижиков В.В., Давенпорт Дж., Чжан К., Ши Е.К., Кабелло О.А., Фукс Дж.Л., Йодер Б.К., Миллен К.Дж .: Белки ресничек контролируют морфогенез мозжечка, способствуя расширению пула предшественников гранул.J Neurosci. 2007, 27: 9780-9789. 10.1523 / JNEUROSCI.5586-06.2007.
CAS Статья Google ученый
Спасский Н., Меркл Ф. Т., Флеймз Н., Трамонтин А. Д., Гарсия-Вердуго Дж. М., Альварес-Буйлла А. Взрослые эпендимные клетки постмитотичны и происходят из радиальных глиальных клеток во время эмбриогенеза. J Neurosci. 2005, 25: 10-18. 10.1523 / JNEUROSCI.1108-04.2005.
CAS Статья Google ученый
Breunig JJ, Sarkisian MR, Arellano JI, Morozov YM, Ayoub AE, Sojitra S, Wang B, Flavell RA, Rakic P, Town T: первичные реснички регулируют нейрогенез гиппокампа, опосредуя звуковые сигналы ежа. Proc Natl Acad Sci USA. 2008, 105: 13127-13132. 10.1073 / pnas.0804558105.
CAS Статья Google ученый
Хан YG, Kim HJ, Dlugosz AA, Ellison DW, Gilbertson RJ, Alvarez-Buylla A: Двойные и противоположные роли первичных ресничек в развитии медуллобластомы.Nat Med. 2009, 15: 1062-1065. 10.1038 / нм.2020.
CAS Статья Google ученый
Амадор-Арджона А., Эллиотт Дж., Миллер А., Гинби А., Пазур Г.Дж., Эниколопов Г., Робертс А.Дж., Терских А.В.: Первичные реснички регулируют пролиферацию усиливающих предшественников в адультиппокампе: значение для обучения и памяти. J Neurosci. 2011, 31: 9933-9944. 10.1523 / JNEUROSCI.1062-11.2011.
CAS Статья Google ученый
Stottmann RW, Tran PV, Turbe-Doan A, Beier DR: Ttc21b необходим для ограничения активности звукового ежа в развивающемся переднем мозге мыши. Dev Biol. 2009, 335: 166-178. 10.1016 / j.ydbio.2009.08.023.
CAS Статья Google ученый
Willaredt MA, Hasenpusch-Theil K, Gardner HA, Kitanovic I, Hirschfeld-Warneken VC, Gojak CP, Gorgas K, Bradford CL, Spatz J, Wolfl S, Theil T., Tucker KL: решающая роль для первичные реснички в корковом морфогенезе.J Neurosci. 2008, 28: 12887-12900. 10.1523 / JNEUROSCI.2084-08.2008.
CAS Статья Google ученый
Kumamoto N, Gu Y, Wang J, Janoschka S, Takemaru K, Levine J, Ge S: роль первичных ресничек в глутаматергической синаптической интеграции нейронов, рожденных взрослыми. Nat Neurosci. 2012, 15: 399-405. 10.1038 / № 3042. S1,
CAS Статья Google ученый
Einstein EB, Patterson CA, Hon BJ, Regan KA, Reddi J, Melnikoff DE, Mateer MJ, Schulz S, Johnson BN, Tallent MK: Передача сигналов соматостатина в ресничках нейронов критична для памяти распознавания объектов. J Neurosci. 2010, 30: 4306-4314. 10.1523 / JNEUROSCI.5295-09.2010.
CAS Статья Google ученый
Отмахова Н.А., Лисман Дж. Э .: Активация дофаминовых рецепторов D1 / D5 увеличивает величину ранней долгосрочной потенциации в синапсах СА1 гиппокампа.J Neurosci. 1996, 16: 7478-7486.
CAS Google ученый
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie.Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie.Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с вашим системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует куки для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
.