Как подводить нижнее веко: 6 главных советов
Часто подводите нижнее веко? У нас есть несколько советов и фотоинструкций, которые помогут улучшить результат
Содержание:
6 советов по макияжу нижнего века
Акцент на нижнее веко – модный прием, который позволяет создать примечательный макияж в прямом смысле одним взмахом руки. Кажется, что проще ничего придумать нельзя, но и в таком мейкапе есть свои нюансы.
Burani © imaxtree
1
Заранее подготовьте «рабочую поверхность»
Если речь идет о подводке слизистой нижнего века, то ее нужно подготовить к нанесению карандаша. Кстати, для такой нежной области следует использовать кайал – особый карандаш с жирной и мягкой текстурой, который не царапает кожу и безвреден для глаз. Прежде чем проводить линию, чистой ватной палочкой пройдитесь по слизистой, чтобы она стала чуть более сухой. По такой поверхности лайнер распределится лучше: цвет будет ярче, а линия – четче.
2
Используйте водостойкие средства
Чтобы не «проморгать» подводку, при работе с нижним веком следует использовать средство с водостойкой формулой. Обычный карандаш, вероятнее всего, ближе к вечеру почти полностью сотрется. Не подведут такие средства как Maybelline New York Expression Kajal и L’Oréal Paris Le Khôl – этим кайалам можете доверить свой макияж.
© Makeup.ru
3
Действуйте по схеме «двойной защиты»
Стойкость подводке нижнего века можно обеспечить и другим способом. После нанесения мягкого кайала возьмите тонкую скошенную кисть. С ее помощью поверх уже проведенной линии распределите немного теней того же оттенка. Этот прием позволит макияжу продержаться гораздо дольше.
4
Держите под рукой карандаши разных оттенков
Черная обводка глаз по контуру действительно может зрительно уменьшить глаза. Чтобы этого не произошло, для нижнего века выбирайте лайнер на пару тонов светлее подводки для верхнего. Например, если стрелки сверху вы рисуете черным, то для нижнего века лучше использовать не угольный кайал, а темно-серый или, например, синий.
5
Обзаведитесь светлым карандашом
Если нижнее веко хочется подвести тем же карандашом, что и верхнее, можно прибегнуть к следующему трюку. Купите белый, нюдовый либо карандаш-кайал в цвете шампанского. После подводки нижнего века основным цветом пройдитесь по слизистой светлым лайнером. Это позволит, во-первых, сделать цвет глаз более ярким, а взгляд – открытым. Во-вторых, избавиться от следов усталости, лицо сделать визуально более бодрым и свежим. Кроме того, после нанесения светлого лайнера темный не будет визуально сужать глаза.
Aquilano e R © imaxtree
China Fashion © imaxtree
6
Не забывайте про растушевку
Визуально глаза станут меньше в том случае, если нижнее веко вы подведете очень четкой, графичной линией. Ключ к наиболее гармоничному макияжу – мягкая линия. После нанесения карандаша слегка растушуйте его подушечками пальцев либо кистью для теней. Благодаря своей мягкой текстуре кайал отлично поддается растушевке.
Elie Saab © imaxtree
Как сделать акцент на нижнем веке: 2 способа
Макияж глаз с точкой на нижнем веке
Burani © imaxtree
Этот небольшой акцент мгновенно преобразит макияж. Понадобятся лишь гелевая подводка и ватная палочка.
Понадобятся лишь гелевая подводка и ватная палочка.
1
Начните с макияжа верхнего века. С помощью жидкой подводки (подойдет L’Oréal Paris Super Liner Superstar) нарисуйте стрелку. Черным карандашом для контура глаз прокрасьте верхнюю слизистую и межресничное пространство. Подчеркните ресницы тушью (мы использовали L’Oréal Paris Volume Million Lashes Fatale).2
По центру нижнего века поставьте точку. Используйте либо черный карандаш, либо гелевую подводку (ее можно нанести ватной палочкой) – такую, как Maybelline New York Lasting Drama Gel Liner.© Makeup.ru
3
Макияж готов!
© Makeup.ru
Макияж нижнего века в технике smoky eyes
Подчеркнуть цвет глаз и создать выразительный макияж поможет один простой трюк – яркий кайал на слизистой нижнего века. Чтобы красиво оттенить природный цвет глаз, выбирайте оттенки, противоположные им по цвету. Для зеленых подойдет карандаш фиолетового или бордового цвета, для голубых – золотой, бронзовый или коричневый, для карих и серый – любой яркий (выбирайте те оттенки, что гармонируют с остальным макияжем).
1
Черным карандашом, например, Slide On Pencil, NYX Professional Makeup, прокрасьте слизистую и межресничное пространство верхнего века.2
Внутренний уголок глаза и подвижное веко подчеркните светлыми сияющими тенями. А внешний уголок глаза и орбитальную линию – матовыми коричневыми (все необходимые цвета есть в палетке NYX Professional Makeup Ultimate в оттенке Warm Rust). Растушуйте с помощью пушистой натуральной кисти.
А внешний уголок глаза и орбитальную линию – матовыми коричневыми (все необходимые цвета есть в палетке NYX Professional Makeup Ultimate в оттенке Warm Rust). Растушуйте с помощью пушистой натуральной кисти.3
Нижнюю слизистую, межресничное и пространство под ресницами закрасьте стойким ярко-фиолетовым карандашом для глаз – таким, как Urban Decay Glide-On 24/7 Eye Pencil в оттенке Psychedelic Sister.© Makeup.ru
4
Используя матовые коричневые тени, растушуйте границы карандаша на нижнем веке. Прокрасьте ресницы черной тушью. Макияж готов!
© Makeup.ru
Фотоидеи для разного цвета глаз
Еще больше идей для макияжа глаз с акцентом на нижем веке ищите в нашей фотогалерее.
Burani © imaxtree
Each x Other © imaxtree
Each x Other © imaxtree
Issa © imaxtree
Еще одну идею макияжа с ярким акцентом на нижнем веке вы найдете в этом видео.
11 идей оригинальных стрелок | BURO.

Макияж глаз в виде тройных стрелок повторил полосатый принт в одежде весенне-летней коллекции Rahul Mishra. Три ярких оттенка подводки нанесены одинаковой ширины линиями от ресничного контура.
Белая подводка выглядит инопланетно и подходит практически всем. Ее можно интересно обыгрывать яркими цветами тушей для ресниц, хотя в образе для показа Laura Biagiotti стилист Aldo Coppola Кьяра Чима использовала классическую черную. Она нарисовала довольно толстые стрелки на верхнем веке и подчеркнула слизистую нижнего.
Не менее авангардный, чем сама коллекция, макияж разработал креативный директор по макияжу Дома Dior Питер Филипс. Центром образа стали сюрреалистичные стрелки с эффектом визуального увеличения размера глаз. Черным лайнером визажист нарисовал линии с ресницами на верхних веках и под нижними.
Кошачьи стрелки могут выглядеть совсем иначе в ярком цвете. Стрелки cat eye должны быть острыми, кончик — вздернутым и далеко выходящим за пределы глаз как во внутреннем уголке, так и во внешнем. Такая стрелка должна растягивать и визуально приподнимать глаза.
Автором этого графичного и контрастного макияжа стал Том Пешо. На подвижное веко визажист нанес тени цвета фуксии, растянув их от внутренних уголков глаз к внешним и вытянув стрелку к вискам. Нижнее веко подведено до середины черным тенями, на верхнем — небольшая стрелка у ресничного контура и более толстая линия, «накрывающая» тени цвета фуксии.
Еще один стильный и необычный прием — стрелка на нижнем веке. Идея была продемонстрирована на показе Michael Sontag в рамках Берлинской недели моды. На внутренних уголках глаз — перламутровые серебряные тени, плавно переходящие в мягкую растушеванную стрелку.
Изящные стрелки, выполненные в технике negative space, несложно повторить самостоятельно. Удобнее всего использовать подводку-фломастер или кисть с плотным ворсом для подводки или с фетровым аппликатором.
Автором футуристичных образов для показа Jeremy Scott осень-зима 2018/19 стал Кабуки. Визажист использовал палетку теней M.A.C. с неоновыми оттенками из новой коллаборации с самим Джереми Скоттом. Разноцветные пластиковые крылья для продолжения стрелок были изготовлены специально для показа.
Разноцветные пластиковые крылья для продолжения стрелок были изготовлены специально для показа.
В создании таких двойных кошачьих стрелок есть нюанс: глаза могут выглядеть слишком суженными. Но дублирование белой подводки вдоль нижнего ресничного ряда визуально увеличивает их и освежает взгляд.
Такие цветные и толстые стрелки можно сделать при помощи плоской широкой кисти и кремовой подводки или теней. Они будут одинаково хорошо смотреться как на открытом веке, так и на нависшем.
Эта идея была представлена на осенне-зимнем показе Prada 2018/19. Автором графичных стрелок стала Пэт МакГрат. Яркими тенями визажист нарисовала широкие стрелки до висков и украсила их кристаллами Swarovski.
Зачем в азиатском макияже стрелка на нижнем веке?: pl_yum — LiveJournal
В XXI веке визажисткий мир разделился на две школы: европейский макияж и азиатский макияж. Причем, азиатский мейк — молодое явление, ибо много лет азиатки пытались вытянуть глаза в традиционную для европейцев миндалевидную форму.
Представьте, насколько удивительно, что сейчас европейки все чаще подсматривают за азиатками и повторяют. Так вот, есть в азиатском макияже прием со стрелокой на нижнем веке, причем эта стрелка смотрит вниз:
А у нас же, у макияжных гур, все вверх, вверх, вверх! Зачем вниз-та?
Следите за руками:
1. Маленький (по европейским меркам) азиатский глаз вытягивается вбок стрелкой на верхнем веке, практически продолжающей разрез
Маленький (по европейским меркам) азиатский глаз вытягивается вбок стрелкой на верхнем веке, практически продолжающей разрез
2. Чтобы достроить форму, добавляем на нижнее веко: усиливаем подводку, дорисовываем разрез и в том числе добавляем стрелку на нижнем веке
Стрелка продолжает разрез глаза, как бы вытягивая его дальше, чем природный разрез. И весь макияж строится вокруг вновь получившегося разреза, а не вокруг настоящего. Так получается новая форма глаза
Стрелка нижнего века, как часть макияжа глаз
Кукольный макияж, в котором изменяется форма нижнего века за счет прорисовки. Видите, тут новый контур прям по коже отрисован.Разве в европейском не так?В европейском мейке стрелка — это повышенная драматичность и декоративный элемент. И стрелка всегда стремится вверх.
Стрелки в азиатском макияже тоже могут смотреть вверх и отвечать той же задача (драма, акцент). А если видите вбок и вниз, то значит таким приемом глаз удлинен (визуально, конечно).
Стрелки на нижнем веке | Салон эстетического татуажа Валиды Мамедовой
Модная тенденция последних двух лет в перманентном макияже – стрелки на нижнем веке. Такие стрелочки нельзя назвать какой-то выделенной самостоятельной техникой, однако они являются отдельной, и очень востребованной услугой в салонах перманентного макияжа.
Особенности техники
Технику прорисовки нижнего века опытные эксперты перманентного макияжа называют «работой без права на ошибку», поскольку именно на нижнем веке исправить любую, даже маленькую неточность в прорисовке стрелки очень сложно, и под силу только высококвалифицированному мастеру.
Набивать нижнюю стрелку, как самостоятельную часть перманентного макияжа глаз, специалисты не рекомендуют. В идеальном варианте – нижние стрелки являются частью локального татуажа век, и наносятся одновременно со стрелками на верхнем веке. Синхронность работы на нижних и верхних веках очень важна, поскольку только при таких условиях вы сможете получить однородный цвет пигмента и оптимальный декоративный рисунок татуажа. В случае если работа будет проводиться разными мастерами или в разное время, высока вероятность расхождений в цвете пигмента и качестве нанесения рисунка, что не только значительно снизит визуальный эффект от работы, но и вынудит вернуться к проблеме ежедневного макияжа, который будет необходим для того, чтобы визуально связать верхнее и нижнее веко в единое цветовое решение.
Особенностью техники нанесения стрелок является ее вариативность – по желанию клиента мастер может вывести классическую стрелку под ресницами, выполнить работу с применением техники мягкой растушевки, либо сделать стрелку в технике межресничного татуажа.
|
Анастасия |
Наталья, Сабина |
Валида |
|
| ПЕРМАНЕНТНЫЙ МАКИЯЖ ВЕК | |||
|
Межресничное пространство |
4.000 |
8.000 |
12.000 |
|
Стрелка классическая |
6.000 |
10.000 |
15.000 |
|
Стрелка с растушевкой |
6. |
10.000 |
15.000 |
|
Нижнее веко |
2.000 |
4.000 |
6.000 |
 000
000
Как выглядит результат?
Визуальный эффект от стрелок на нижнем веке виден сразу – красиво подчеркивается форма глаз, в образе проявляется «восточный» стиль и яркий акцент. При правильном выборе дизайна нижних стрелок, взгляд становится более глубоким выразительным, визуально увеличивается разрез и форма глаз.
Кожа на нижнем веке особо чувствительна к взаимодействию с пигментом, поэтому результат, полученный во время процедуры, будет долго радовать обладательницу стрелок четким и ярким рисунком без каких либо дополнительных усилий.
Для кого?
Татуаж нижних век универсален. Он является дополнением к верхним стрелкам, и подходит женщинам с любым разрезом глаз и типом лица и кожи. Особенно актуален для обладательниц миндалевидных глаз и женщин, предпочитающих «восточный» стиль в декоративном макияже.
Как правильно подводить нижнее веко карандашом: пошаговые уроки с фото
Один из способов сделать в макияже яркий акцент – подчеркнуть цветом линию под нижними ресницами. Важно сделать это правильно, чтобы не утяжелить взгляд и визуально не уменьшить глаза. Сегодня я дам вам в помощь несколько лайфхаков, расскажу подробнее о карандашах для глаз и покажу простой пошаговый урок с фото.
6 хитростей макияжа с карандашной подводкой
Сама по себе техника очень проста: достаточно взять лайнер нужного цвета и провести линию. Здесь сложно ошибиться с формой: это не сложная стрелка вдоль верхних ресниц. Но есть несколько профессиональных хитростей, которые помогут вам сделать подводку по нижнему веку еще более красивой и стойкой.
Давайте разберемся, как правильно подводить нижнее веко:
- Перед нанесением проведите по слизистой ватной палочкой.
 Это нужно, чтобы сделать поверхность суше, и нарисованная линия держалась дольше. Кроме того, так подводка будет более насыщенной по цвету.
Это нужно, чтобы сделать поверхность суше, и нарисованная линия держалась дольше. Кроме того, так подводка будет более насыщенной по цвету. - Закрепляйте подводку для большей стойкости. Для этой цели идеально подходят сухие тени близкого оттенка: наберите немного на пушистую кисть и нанесите поверх карандашной линии.
- Если не хотите визуально уменьшить глаза, наносите на слизистую только светлый лайнер. Такой прием сделает взгляд более открытым за счет того, что вы сотрете нежелательный контраст между белком глаза и веком.
 Это нужно, чтобы сделать поверхность суше, и нарисованная линия держалась дольше. Кроме того, так подводка будет более насыщенной по цвету.
Это нужно, чтобы сделать поверхность суше, и нарисованная линия держалась дольше. Кроме того, так подводка будет более насыщенной по цвету.- Комбинируйте стрелки разных оттенков. Только черная подводка не так интересна, как игра цвета. Например, если на верхнем веке у вас классическая стрелка, вдоль нижних ресниц проведите серую или яркую цветную линию.
- Стремитесь к мягким линиям. Графичная, четкая подводка также способна уменьшить глаза. Чтобы этого не допустить, всегда держите под рукой синтетическую кисточку и тщательно растушевывайте подводку. Также это можно сделать подушечками безымянных пальцев.
- Вместо полноценной стрелки можно рисовать уголок – не более 1/3 вашего глаза. Чтобы подводка по линии нижних ресниц выглядела более мягко и легче сочеталась с другими яркими акцентами, просто не дорисовывайте стрелку до конца.
Все эти приемы используются на подготовленном веке. Прежде чем подвести веко карандашом, нужно нанести консилер, немного пудры и базу для макияжа глаз. Так линия получится более стойкой и яркой.
Какие карандаши для глаз удобно иметь в косметичке?
Подводка по линии нижних ресниц может быть разной – нейтральной, черной или цветной. Вы можете провести линию прямо по слизистой или немного отступить и сделать мягкую тушевку. И для каждой задачи нужно свое средство.
Чтобы свободно создавать вечерние и дневные образы, вам понадобятся такие продукты:
- Мягкий кайал для слизистой. Кайалом называется карандаш с нежной, чуть более жирной текстурой. Он оставляет на слизистой оболочке века плотную линию с первого раза и очень комфортно наносится. Если вы хотите использовать светлый кайал, выберите вместо белого более естественный оттенок – нежно-розовый, бежевый или персиковый.
- Черный или темно-коричневый лайнер для традиционной подводки по нижнему веку. Пригодится вам и в вечернем, и в дневном макияже: можно подвести нижнее веко карандашом такого цвета в соседстве с любыми тенями. Чтобы линия не выглядела грубо, обязательно растушуйте ее чистой кистью.
- Яркие акцентные варианты: изумрудный, синий, лиловый и т.д. Контрастная линия вдоль века будет привлекать внимание и выгодно оттенять ваши глаза. Кареглазым девушкам пойдут насыщенные темные оттенки, а если у вас серая или голубая радужка, выбирайте более светлые тона.
 Он оставляет на слизистой оболочке века плотную линию с первого раза и очень комфортно наносится. Если вы хотите использовать светлый кайал, выберите вместо белого более естественный оттенок – нежно-розовый, бежевый или персиковый.
Он оставляет на слизистой оболочке века плотную линию с первого раза и очень комфортно наносится. Если вы хотите использовать светлый кайал, выберите вместо белого более естественный оттенок – нежно-розовый, бежевый или персиковый.Независимо от цвета и текстуры, лучше выбирать водостойкие лайнеры. Средство с такой формулой гарантированно останется на месте до самого вечера и легко смоется специальной жидкостью. Это особенно важно для кайала, который наносится прямо на слизистую.
Простой пример макияжа с акцентом на нижнем веке
Мы разобрались в тонкостях процесса и отобрали несколько подходящих средств – теперь посмотрим на примере, как подводить нижнее веко карандашом. Предлагаю несложный мейк-ап с использованием белого, синего и черного лайнера. Помимо теней и подводки вам потребуется скошенная кисть.
В каком порядке создается такой образ:
- Растушуйте белый или розоватый кайал. Используйте его вместо кремовых теней и нанесите на верхнее веко, в уголок глаза и на слизистую. Сделать покрытие однородным вам поможет кисть или подушечки пальцев.
- Сделайте яркий акцент по линии нижних ресниц. Для этого вам понадобится мягкий лайнер голубого, темно-синего или бирюзового оттенка. Просто нарисуйте стрелку – растушевывать ее мы будем следующим шагом.
- Смягчите линию и закрепите стрелку. Теперь возьмите жесткую скошенную кисть и хорошенько растушуйте линию, чтобы смягчить эффект и сделать взгляд легче. После этого наберите на кисть немного синих теней и покройте ими свежий слой подводки. Чтобы понять принцип, посмотрите, как подведено карандашом нижнее веко на фото.
 Теперь возьмите жесткую скошенную кисть и хорошенько растушуйте линию, чтобы смягчить эффект и сделать взгляд легче. После этого наберите на кисть немного синих теней и покройте ими свежий слой подводки. Чтобы понять принцип, посмотрите, как подведено карандашом нижнее веко на фото.
Теперь возьмите жесткую скошенную кисть и хорошенько растушуйте линию, чтобы смягчить эффект и сделать взгляд легче. После этого наберите на кисть немного синих теней и покройте ими свежий слой подводки. Чтобы понять принцип, посмотрите, как подведено карандашом нижнее веко на фото.- Завершите образ классической черной стрелкой. На этот раз линию нужно провести вдоль верхних ресниц – удобно использовать гелевую подводку и наносить ее так же кистью. Чтобы получить более эффектный макияж, заведите немного черного и на нижнее веко: сформируйте уголок стрелки. Рисуйте прямо по синему лайнеру.
Всего за 4 шага макияж готов. Остается дополнить его тушью и немного подкрасить брови и губы.
10 модных макияжей для этой весны — Wonderzine
Полгода назад визажисты на показах работали не покладая рук над тем, чтобы предложить нам свежие идеи макияжа для этой весны. Мы выбрали самые классные образы, которые рекомендуем примерить с наступлением тепла (или до него).
Rochas
Нам, определенно, нравится тренд на отсутствие перфекционизма, который прослеживался на некоторых шоу. Так, для Louis Vuitton визажисты обвели глаза «ежиком», а для Rochas прямо ватной палочкой поставили на нижнем веке жирные черные точки — их сочетание со склеенными ресницами и светло-голубыми тенями получилось трогательным, небанальным и, что самое приятное, простым в исполнении.
Zac Posen
Красная помада никогда не сойдет с подиумов — ну, только если визажисты не организуют заговор против нее. Мы отлично понимаем ее плюсы и минусы (всегда красиво, но может надоесть) и не призываем никого отказываться от классических образов. Тем не менее нам нравится разбавлять привычный образ — например так, как это сделали визажисты для Zac Posen. Блеск на веках не самое привычное ощущение — зато это один из самых быстрых способов накрасить глаза так, чтобы они ловили взгляды.
Givenchy
Показ Givenchy стал известен многим из-за того, что СМИ растиражировали его как «первый выход Кары Делевинь без бровей». Меж тем интересен он не этим: выбеленными бровями мало кого можно сейчас удивить, а вот охристые стрелки запомнились тем, что придали моделям хищные и одновременно птичьи черты. Не обязательно повторять макияж в точности (если достаточно смелости, почему бы и нет), но можно хотя бы пересмотреть привычные схемы нанесения цветных теней.
Меж тем интересен он не этим: выбеленными бровями мало кого можно сейчас удивить, а вот охристые стрелки запомнились тем, что придали моделям хищные и одновременно птичьи черты. Не обязательно повторять макияж в точности (если достаточно смелости, почему бы и нет), но можно хотя бы пересмотреть привычные схемы нанесения цветных теней.
Elie Saab
Макияж Elie Saab с его выделенными бровями и темно-серыми смоки был бы скучен, не используй визажисты вместо черного кайала ярко-голубой. Этот трюк сработал бы и с карандашами любого другого цвета, так что если вы уже научились растушевывать серые тени, то с лайфхаком Elie Saab можно открыть для себя дивный новый мир макияжа глаз и быстро краситься по-новому хоть каждый день.
Just Cavalli
Just Cavalli тоже поддержали тренд на несерьезность и не стали выводить предельно аккуратные стрелки, а просто подвели нижнее веко так, что на большее не хватило времени. Да, это макияж не из разряда общепринято привлекательных, но кто сказал, что именно к этому нужно всегда стремиться?
Ashish
На моделей Ashish, кажется, налепили всё лучшее сразу: жемчужины, звёзды, стразы и много-много глиттера — даже на ногтях. Мы не раз признавались в любви к макияжам с разными наклейками, и этот дискотечный вариант Ashish нам особенно импонирует. Да, в большинство офисов так не пойдешь, но почему бы вместо черных стрелок не приклеить себе маленькие звёзды у внешних уголков глаз? Едва ли сможем придумать более простой и занятный выбор на каждый день.
Preen
Веснушки не только не недостаток (не можем понять, почему некоторые их обладатели стесняются), но украшение — это решили продемонстрировать визажисты сразу нескольких показов. Нет ничего удивительного, что в шоу Vivienne Westwood их изобразили театрально, но вариант, показанный Preen, нам кажется абсолютно уместным в повседневности. Да, можно не расставлять веснушки по всему лицу, а ограничиться, например, скулами и носом (кстати, использовать для этого можно обычный светло-коричневый карандаш для глаз).
Holly Fulton
Еще один очень простой и почти гениальный трюк: не подводить слизистую одним цветом, а скомбинировать два. Голубой карандаш придает взгляду свежести, а белый, как мы помним, маскирует естественные покраснения — а если он еще и такой яркий, то дополнительно выделяет глаза. Если бы нас попросили назвать идеальный летний макияж, то у такого были бы все шансы победить.
Dior
Не могли обойти вниманием очередную вариацию макияжа с наклейками. Если в только что ушедшую зиму Питер Филипс предлагал носить на глазах металл, то весной и летом его подменят тканевые стрелки пастельных оттенков. Тем, кто хочет скопировать образ, марка дает возможность купить те самые стрелки Dior Pastel Eyes, но можно обойтись и малыми средствами — мы вот любим клеить на себя всё, что не вредно, и замену фирменным стрелкам усматриваем в обычной плотной ткани.
Tommy Hilfiger
Закроем тему простого и эффектного макияжа работой Tommy Hilfiger: здесь опять натуральный макияж, но с веселым акцентом в виде фигурных пайеток. Визажисты клеили их не только на лицо, но и на область декольте, и это еще раз доказывает, что для макияжа нет границ даже географических. Если соберетесь воспроизвести что-то подобное, приклеить украшения можно обычным клеем для накладных ресниц.
ФОТОГРАФИИ: Getty Images/Fotobank (10)
Стрелка нижнего века, татуаж стрелки на нижнем веке в Москве
Довольно оригинальным видом перманентного макияжа является стрелка нижнего века. Для тех, кто привык подводить глаза карандашом, в том числе и под нижними ресницами, такая процедура подарит возможность надолго забыть о декоративной косметике.
Стрелка на нижнем веке, созданная опытными мастерами, выглядит аккуратно и элегантно. Сделав татуаж, вы сможете выглядеть идеально в любую погоду, независимо от ситуации, а ваш выразительный взгляд непременно привлечет внимание окружающих.
Перманентный макияж нижнего века — довольно эффектный вариант татуажа глаз. Однако стрелки нижнего века визуально сужают глаза и делают их меньше. Поэтому стоит учесть, что такой татуаж подойдет далеко не каждой женщине. Процедура нанесения красящего пигмента на кожу под нижние ресницы актуально для обладательниц больших, распахнутых глаз, чтобы подчеркнуть их выразительность.
Однако стрелки нижнего века визуально сужают глаза и делают их меньше. Поэтому стоит учесть, что такой татуаж подойдет далеко не каждой женщине. Процедура нанесения красящего пигмента на кожу под нижние ресницы актуально для обладательниц больших, распахнутых глаз, чтобы подчеркнуть их выразительность.
Современные техники нанесения татуажа на нижнее веко:
- Прорисовка межресничного пространства. Такой вариант макияжа позволит вам выглядеть максимально естественно, словно вы ненакрашены. После процедуры ваш взгляд станет глубже, а глаза покажутся еще ярче.
- Четкая стрелка. Данный вид татуажа представляет собой нанесение на кожу тонкого сплошного контура вдоль линии роста нижних ресниц.
- Татуаж с растушевкой. Вариант, представляющий собой нанесение краски с размытыми краями для создания эффекта теней. Подходит также для восстановления и исправления ранее нанесенного неровного контура.
Татуаж нижнего века в центре «Де Маре»
Мастера центра перманентного макияжа «Де Маре» не только качественно и безболезненно сделают татуаж, но и помогут выбрать вид стрелок, подходящий к форме ваших глаз. Процедура производится на современном оборудовании с использованием высококачественных красящих пигментов. Вы можете выбрать как классический черный цвет, так и более мягкие, светлые оттенки для нанесения татуажа.
Процесс заживления после нанесения перманентного макияжа под нижние ресницы практически безболезнен. После завершения самой процедуры наши мастера дадут все необходимые рекомендации по уходу за татуажем, чтобы вы смогли как можно быстрее увидеть окончательный результат.
Крайне важно в первые дни после процедуры нанесения стрелок на нижнее веко использовать заживляющую мазь и защищать появляющиеся корочки от воды, косметических средств и солнечных лучей. При соблюдении всех правил по уходу заживление займет не более двух недель, а качественный татуаж будет радовать вас на протяжении трех лет и более.
Заболевания век | Институт офтальмологии
В ESI мы лечим многие заболевания век. Общие условия включают:
Общие условия включают:
Блефарит
Это очень распространенное постоянное состояние краев век обоих глаз, при котором воспаление сальных желез на верхнем и нижнем веках вызывает покраснение, зуд и симптомы сухости глаз. Это связано с кожными бактериями, розацеа лица и, в некоторых случаях, заражением демодексом. К сожалению, лечение этого состояния требует регулярной чистки век / гигиенических методов для минимизации симптомов.Другие стандартные процедуры включают массаж век, теплые компрессы, смазку глаз, кремы для глаз с антибиотиками и стероидами, а иногда и пероральные антибиотики. Для лечения демодекса используются препараты масла чайного дерева (50% раствор), появляются новые методы лечения, в том числе паровые и лазерные. Дополнительную информацию об этом состоянии можно получить на приеме у специалиста или в клинике по лечению сухого глаза.
Стрелки на нижнем веке показывают воспаление мейбомиевых сальных желез
Стрелки на верхнем веке показывают перхоть вокруг основания ресницы
Халязион / ячмень / киста века
Ячмень и халазия — это кисты или уплотнения век, которые могут возникнуть в любом возрасте.Часто они связаны с основным блефаритом. Они представляют собой механическую блокировку сальных желез на веке и обычно не вызваны инфекцией. Они могут вызывать неприятное покраснение, отек и боль века, но обычно проходят самостоятельно.
Большая часть опухолей будет набухать до тех пор, пока киста не станет точками и содержимое кисты не выйдет через кожу века или изнутри. Применение горячих компрессов в течение 15 минут 4 раза в день в течение 5 дней может ускорить этот процесс:
- используйте кипяченую воду (не ошпаривая) на чистой фланели или марле
- или нагрейте металлической ложкой, обернутой марлей, и прижмите к пораженному веку
Иногда халязион прогрессирует, вызывая инфекцию века и ассоциируется с усилением боли, прогрессирующим покраснением кожи века и усилением отека. В этом случае важно начать прием антибиотиков перорально или внутривенно, и вам следует немедленно обратиться за помощью.
В этом случае важно начать прием антибиотиков перорально или внутривенно, и вам следует немедленно обратиться за помощью.
Иногда киста халязиона не рассасывается, и внутри века остается небольшая остаточная плотная шишка. В этом случае киста потребует рассечения и дренирования, чтобы ускорить ее рассасывание. Ваш специалист может дать вам соответствующий совет.
Опухоли / рак кожи
По статистике, люди, живущие в Квинсленде, имеют самый высокий в мире шанс заболеть раком кожи.Примерно 50% случаев рака кожи обнаруживаются на лице или шее из-за воздействия солнца, и многие из них находятся в области век. Опухоли кожи могут быть доброкачественными (нераспространяющиеся новообразования) или злокачественными (злокачественными). Большинство небольших бугорков на веках безвредны, но вам всегда следует обращаться за консультацией к офтальмологу или кожному специалисту.
Среди видов рака кожи преобладают немеланомные виды рака кожи, причем наиболее распространенным является базально-клеточная карцинома (BCC). Обычно это медленно растущая опухоль, распространение которой за пределы исходного места роста встречается крайне редко.Другие виды рака век включают плоскоклеточную карциному (SCC) и карциному сальных клеток. Они обычно имеют более быстрый образец роста и могут распространяться за пределы исходного места роста. Меланома также может возникать вокруг периокулярной области.
Каковы симптомы рака век?
Рак век обычно начинается с небольшого безболезненного поражения кожи, которое является твердым или узловатым и продолжает расти. Они могут стать твердыми и казаться прикрепленными к более глубоким тканям. Иногда они будут кровоточить, изъязвляться или быть коричневыми (пигментированными).Шишки, появляющиеся на веке, могут доходить до края века или затрагивать конъюнктиву с не устраняемым покраснением края века. Обратитесь к своему специалисту, если у вас есть наросты, которые демонстрируют эти особенности.
Как диагностируется рак век?
Большинство поражений будут иметь типичный вид для глазного специалиста. Однако в условиях клиники может потребоваться небольшая биопсия (пункционная биопсия) ткани под местной анестезией, чтобы обезболить пораженную область, чтобы обеспечить надлежащее планирование наилучшего лечения опухоли.Иногда ваш специалист запросит МРТ-сканирование этой области, чтобы оценить глубину опухоли и ее отношение к связанным тканям.
Однако в условиях клиники может потребоваться небольшая биопсия (пункционная биопсия) ткани под местной анестезией, чтобы обезболить пораженную область, чтобы обеспечить надлежащее планирование наилучшего лечения опухоли.Иногда ваш специалист запросит МРТ-сканирование этой области, чтобы оценить глубину опухоли и ее отношение к связанным тканям.
Как лечится рак век?
Тщательное рассмотрение механики и функции век имеет важное значение для наилучшего лечения периокулярных новообразований. Доступны различные варианты лечения, но обычно предпочтительнее хирургическая резекция.
В качестве офтальмолога ваш консультант в ESI сможет тщательно сбалансировать удаление опухоли с сохранением функции вашего века и защитой поверхности глаза.Небольшие поражения можно удалить в палатах клиники под местной анестезией. В противном случае удаление может быть выполнено в условиях театра с рядом доступных вариантов анестезии.
Неправильное положение нижнего века
Положение и тон нижнего века важны для нормальной защиты глаз и оттока слез. Со временем, предыдущей операцией или воспалительными состояниями кожи или конъюнктивы, веки могут стать перевернутыми (эктропион) или перевернутыми (заворот).Таким образом, защита глазной поверхности и система отвода слез могут быть нарушены.
Каковы симптомы неправильного положения нижних век?
Энтропион или эктропион проявляются симптомами покраснения глаз и век, болью в глазах, ощущением инородного тела, рецидивирующими инфекциями / конъюнктивитом, слезотечением и ухудшением зрения. Внешний вид перевернутого века может быть очевидным, но часто видны лишь незначительные изменения, а симптомы заболевания.
Ectropion — веко откидывающееся
Entropion — нижнее веко закатано внутрь
Как лечится неправильное положение нижнего века?
Для устранения проблемы необходимо устранить конкретную причину и защитить глаза.
- Временные меры могут включать смазывающие глазные капли и капли с антибиотиками или мази от рецидивирующих инфекций. Для энтропиона (обратного хода) в некоторых случаях может быть сделана инъекция миорелаксанта, что даст примерно 3 месяца улучшения. Кроме того, липкую ленту можно наложить от нижнего века на щеку, чтобы временно оттянуть нижнее веко от глаза и облегчить симптомы.
- Постоянное лечение обычно хирургическое.
Каковы риски операции?
Как и при любой хирургии век, существует небольшой риск образования рубцов, инфекции, синяков, необходимости в дополнительной операции, нечеткости зрения или трудностей с закрытием век. Ваш специалист проконсультирует вас.
Чего мне ожидать во время и после операции?
Операция на нижнем веке обычно проводится под местной анестезией в дневном стационаре. Седация может помочь вам меньше осознавать операцию и уменьшить беспокойство.Также доступен общий наркоз (для полного сна). Ваш хирург может попросить вас скорректировать ваши обычные разжижающие кровь средства перед операцией, если вы их принимаете, и лучше всего избегать рыбьего жира и чеснока в течение 2 недель до операции.
После операции вы увидите своего специалиста примерно через 1 неделю, чтобы снять швы и оценить заживление раны. Внешний вид нижнего века может казаться слегка высоким и болезненным в течение 6 недель после операции, но затем обычно полностью оседает. Вам дадут крем с антибиотиком для втирания в любые раны век, который безопасен для глаз.
Что вызывает темные круги под глазами? — Адам Шайнер MD
С возрастом мы обнаруживаем, что одно из самых заметных изменений нашей внешности касается области вокруг глаз . Пожалуй, самая частая жалоба, которую я получаю как специалист по косметическому омоложению век, заключается в том, что мои пациенты слышали от других, что они выглядят больными или усталыми, даже когда чувствуют себя хорошо и отдохнувшими.
Источником таких нежелательных комментариев обычно является невербальное сообщение , которое область нижнего века отправляет по мере того, как мы стареем на год. На эту область может влиять множество различных факторов, каждый из которых способствует появлению темных кругов под глазами. Первый — это выпуклость или вогнутость жировой ткани нижнего века. Если жир нижнего века выступает в область нижнего века , он часто улавливает тени над головой.
На эту область может влиять множество различных факторов, каждый из которых способствует появлению темных кругов под глазами. Первый — это выпуклость или вогнутость жировой ткани нижнего века. Если жир нижнего века выступает в область нижнего века , он часто улавливает тени над головой.
Взгляните на тени, показанные синими стрелками на фотографии этого пациента, которому я помог с операцией по омоложению нижних век.
Поскольку большая часть света находится над нами в нашем мире (рассмотрим солнечный свет снаружи и комнатное освещение внутри), этот свет, вероятно, отбрасывает тень на выступающий жир, который становится видимым на коже под жиром нижнего века. Результатом затенения являются непривлекательные темные круги под глазами, сигнализирующие об усталости или болезни.
Обратная проблема, связанная с жиром нижнего века, встречается редко, но может возникать изредка и включает недостаток жира в области нижнего века , вызывая появление затонувшего века, которое аналогичным образом улавливает и отображает тени на коже нижнего века. Это вторая причина появления темных кругов под глазами и появления темных кругов на глазах.
Третий фактор, способствующий появлению темных кругов под глазами, — это темный пигмент , который может откладываться на коже в области нижнего века.Обычно это результат пребывания на солнце в течение всей жизни.
Обратите внимание на фотографию этого пациента перед операцией, у которого были множественные темные возрастные пятна и пигмент на коже нижнего века, показанные синими стрелками. Эти пятна, а также внешний вид верхнего и нижнего века мне помогли с помощью моих техник омоложения верхнего и нижнего века.
Четвертый фактор, который испытывают пациенты, приводящий к темным кругам под глазами, — это нижележащих сосудов, проходящих через тонкую кожу в области нижнего века. Это видно на фотографии пациента, у которого были две причины появления темных кругов под глазами. У них были тени, улавливающие полноту, но, как демонстрирует эта фотография, у них также была тонкая кожа, показывающая розовые сосуды в этой области кожи нижнего века ближе к носу.
Это видно на фотографии пациента, у которого были две причины появления темных кругов под глазами. У них были тени, улавливающие полноту, но, как демонстрирует эта фотография, у них также была тонкая кожа, показывающая розовые сосуды в этой области кожи нижнего века ближе к носу.
Наконец, пятый фактор, который может играть роль в появлении темных кругов под глазами, — это наличие набухания нижнего века и щек , известных как нижнее веко, фестоны на щеках и скуловые бугры.Это видно по синим стрелкам на фотографии пациентки, указывающей на фестоны до того, как я помог ей с процедурами омоложения нижних век.
Вот пять наиболее частых причин появления темных кругов под глазами:
- Полнота под глазами улавливаются тени.
- Впадина Под глазами улавливаются тени.
- Коричневый пигмент или пигментные пятна на коже под глазами.
- Тонкая кожа и покраснение, проступающее сквозь кожу в области под глазами.
- Festoons и скуловые бугры, улавливающие тени в области нижних век и щек., / Li>
Хотя эти пять распространенных причин темных кругов под глазами могут приводить к неоптимальным невербальным сообщениям, существуют варианты их лечения , омолаживая внешний вид и восстанавливая эффективное невербальное общение .
Помогать моим пациентам с этими проблемами — настоящее удовольствие.Я испытываю огромное удовлетворение, когда пациенты сообщают мне после того, как я помогаю им, что другие люди в мире лучше относятся к ним, потому что эти различные причины темных кругов исчезли.
Для получения дополнительной информации о косметическом омоложении век, фестонах, темных кругах под глазами или услугах косметической хирургии Тампа, включая блефаропластику и ботокс, свяжитесь с нашим офисом.
Нижняя блефаропластика — д-р Ли Нижний
Блефаропластика, также известная как хирургия век, — это хирургическая процедура, которая улучшает внешний вид век путем подтягивания мышц и тканей или удаления излишков жира и кожи. Блефаропластика нижнего века используется для лечения:
- Мешки под глазами
- Опущенные нижние веки
- Лишняя кожа
- Мелкие морщинки
- Круги под глазами
Блефаропластика нижнего века может быть сделана как взрослым мужчинам, так и женщинам, она обеспечивает более молодой и свежий вид, который отражается на всем лице.
Блефаропластика — одна из наиболее часто выполняемых процедур пластической хирургии лица.Популярность этой процедуры отражает важность глаз в улучшении внешнего вида. Если внешний вид глаз вызывает у пациента недовольство, он может рассмотреть возможность блефаропластики нижнего века, чтобы улучшить внешний вид и повысить уверенность в себе с помощью безопасной процедуры с минимальным временем простоя.
Кандидаты на блефаропластику нижнего века
Лучшими кандидатами на блефаропластику нижнего века являются лица, отвечающие следующим условиям:
- Хорошее общее состояние здоровья
- Не курить
- Нет серьезных заболеваний глаз
- Иметь здоровые ткани и мышцы лица
Если у пациента дряблая, дряблая кожа или опухшие мешки под глазами, эта процедура может помочь уменьшить эти нежелательные эффекты и сделать глаза молодыми и свежими.
Важно, чтобы пациенты имели реалистичные ожидания перед процедурой блефаропластики нижнего века, чтобы они остались довольны своими результатами. Хотя процедура может улучшить их внешний вид и в результате повысить уверенность в себе, она не может полностью изменить внешний вид пациента или структуру его лица.
Процедура блефаропластики нижнего века
Блефаропластика нижнего века проводится под местной или общей анестезией.Используемый метод или процедура будет зависеть от количества жира, который необходимо удалить, расположения жира и того, насколько плотно сделать веко. Процедура обычно занимает от 45 минут до часа.
Существует несколько различных хирургических подходов, которые можно использовать при блефаропластике нижнего века. Они следующие:
Они следующие:
Трансконъюнктивальная блефаропластика нижнего века
Трансконъюнктивальная блефаропластика нижнего века, которую часто выбирают, когда не нужно удалять кожу, создает разрез на внутренней стороне нижнего века, не оставляя видимых шрамов или других следов операции.
Чрескожная блефаропластика нижнего века
Чрескожная блефаропластика нижнего века выполняется с помощью субцилиарного разреза, также известного как щипковая техника, для удаления выступающего жира и лишней кожи.
Трансконъюнктивальный Arcus Marginalis, выпуск
Освобождение трансконъюнктивальной дуги marginalis перемещает жир внутри века для поддержания некоторого объема в мягких тканях. Это предотвращает чрезмерное выпадение гребня орбитальной кости, из-за чего глаза выглядят пустыми, а лицо — старше.
После выполнения процедуры блефаропластики нижнего века разрезы зашивают швами и обычно оборачивают марлей для заживления области.
Восстановление после блефаропластики нижнего века
После процедуры блефаропластики нижнего века хирург может порекомендовать применять смазывающие капли или мазь и холодные компрессы, чтобы помочь в процессе заживления и минимизировать побочные эффекты, такие как:
- Набухание
- Ушиб
- Раздражение
- Сухие глаза
Глаза также можно закрыть марлей после процедуры.Пациенты обычно могут вернуться к работе в течение нескольких дней, но им следует избегать физических упражнений и физических нагрузок в течение как минимум двух недель. Швы обычно снимают через три-четыре дня после процедуры. Отеки и другие побочные эффекты обычно проходят в течение двух недель после операции. Контактные линзы и макияж век нельзя носить в течение двух недель после операции. Хирург даст конкретные инструкции о том, как ухаживать за глазами после процедуры.
Результаты блефаропластики нижнего века
Результаты блефаропластики нижнего века видны по мере исчезновения отека и синяков, которые будут улучшаться в течение года после операции. Процедура блефаропластики нижнего века предназначена для длительного результата. Хотя операция не может предотвратить старение глаз, большинство пациентов довольны своими результатами и не стремятся к повторной процедуре.
Процедура блефаропластики нижнего века предназначена для длительного результата. Хотя операция не может предотвратить старение глаз, большинство пациентов довольны своими результатами и не стремятся к повторной процедуре.
Риски и осложнения блефаропластики нижнего века
Как и любая хирургическая процедура, блефаропластика нижнего века сопряжена с определенными рисками. Эти риски редки, но включают следующее:
- Ушиб
- Набухание
- Расплывчатое зрение
- Неправильное положение глаза
- Кровотечение
- Инфекция
- Сухие глаза
- Изменение цвета кожи
- Невозможность полностью закрыть глаза
Дополнительные ресурсы
процедур для век Нортгемптон | Хирургия век Springfield
Мы знаем веко
Балинский глазной и лазерный центр — это комплексный центр лечения глаз, специализирующийся как на состоянии глаз, так и на веках.Доктор Балин выполняет косметическую операцию на веках для лечения аномалий век, вызванных болезнью, травмой или естественным старением. Доктор Балин регулярно удаляет кисты, новообразования и раковые образования в области вокруг глаз. Она также выполняет блефаропластику для лечения лишней кожи век, а также операции по поводу птоза, эктропиона и энтропиона.
Энтропион и эктропион могут вызывать у пациентов болезненные симптомы. Избыточная кожа век может ограничивать поле зрения пациента и вызывать некоторые неблагоприятные симптомы. Хирургия век может уменьшить болезненные симптомы этих состояний, а также улучшить поле зрения пациента и общий вид глаза.Перед операцией тщательно исследуют глаз и веко. Операция проводится офтальмологом доктором Балином в нашем хирургическом кабинете, расположенном в Нортгемптоне, в глазном и лазерном центре Балина в амбулаторных условиях с использованием местной анестезии.
Птоз — опущение верхнего века
Птоз — это состояние, при котором верхнее веко существенно опускается над глазом, тем самым ограничивая поле зрения. Это состояние может быть очевидным при рождении (врожденное) или развиваться с возрастом (инволюционное).
Это состояние может быть очевидным при рождении (врожденное) или развиваться с возрастом (инволюционное).
Все дети с врожденным птозом должны быть обследованы офтальмологом или глазным хирургом, чтобы убедиться в отсутствии других заболеваний глаз. В дошкольном возрасте рекомендуется проводить операцию, чтобы ребенку было легче видеть и улучшить его внешний вид. Фактически выполняемая операция зависит от тяжести состояния.
Инволюционный птоз развивается с возрастом и может ухудшиться после операции на глазах или отека век. Опущенное веко может ограничить поле зрения и придать глазам неровный вид.Хирург-офтальмолог может укоротить мышцу, открывающую веко, чтобы вылечить это состояние.
Избыточная кожа век
По мере естественного старения кожа теряет эластичность. Поскольку кожа век — самая тонкая кожа на теле, она очень чувствительна к растяжению, вызывая обвисание верхнего века и мешки на нижнем веке. Этот избыток кожи влияет на наш общий внешний вид, а также ограничивает поле зрения. Доктор Балин может провести операцию на глазах в нашем офисе в Нортгемптоне. Удаляется лишняя кожа, а также любая жировая ткань, которая может присутствовать, чтобы вернуть векам их нормальное положение.
Эктропион — поворот нижнего века наружу
Чрезмерное растяжение кожи нижнего века может привести к опусканию века и повороту наружу. Это также может произойти от ожогов век или некоторых кожных заболеваний. Это состояние может вызвать сухость глаз, чрезмерное слезотечение, покраснение и чувствительность к свету и ветру. Эти симптомы можно уменьшить, вернув нижнее веко хирургическим путем в исходное положение.
Энтропион — поворот нижнего века внутрь
Чрезмерное растяжение нижнего века может привести к опусканию века и повороту внутрь.Это состояние также может быть вызвано инфекцией и рубцами внутри века. Когда веко поворачивается внутрь, ресницы и кожа раздражают глаз, постоянно трясь о них. Это приводит к тому, что глаза становятся красными и чувствительными к свету и ветру. Если не лечить Энтропион, он может привести к образованию язвы глаза. Чтобы вернуть веко в нормальное положение, необходима операция.
Если не лечить Энтропион, он может привести к образованию язвы глаза. Чтобы вернуть веко в нормальное положение, необходима операция.
Произошла ошибка при настройке вашего пользовательского файла cookie
Произошла ошибка при настройке вашего пользовательского файла cookieЭтот сайт использует файлы cookie для повышения производительности.Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
FGF-регулируемая передача сигналов BMP необходима для закрытия век и для определения судьбы эпителиальных клеток конъюнктивы
Резюме
Имеются противоречивые отчеты о том, требуется ли передача сигналов BMP для
закрытие век при внутриутробном развитии плода. Этот вопрос был решен с использованием
мыши, дефицитные по передаче сигналов BMP или TGFβ в проспективном веке и
эпителиальные клетки конъюнктивы. Гены, кодирующие два рецептора BMP типа I,
рецептор TGFβ типа II, два BMP- или два активированных TGFβ R-Smads или
co-Smad Smad4 были удалены из эктодермы поверхности глаза с использованием Cre
рекомбиназа.Только мыши с делецией компонентов пути BMP имели
Фенотип «открытое при рождении веко». Мыши без Fgf10 или Fgfr2 также имеют открытые веки при рождении. Чтобы лучше понять
пути, которые регулируют экспрессию и функцию BMP во время развития век,
мы локализовали BMP и промежуточные соединения передачи сигналов BMP в Fgfr2 и Smad4 мышей с условным нокаутом (CKO). Мы обнаружили, что Fgfr2 был
требуется для выражения Bmp4 , нормальное распределение Shh
сигнализация и для сохранения дифференцировки конъюнктивы
эпителий.Передача сигналов FGF также способствовала экспрессии антагониста Wnt.
Sfrp1 и подавление передачи сигналов Wnt в предполагаемых эпителиальных клетках век,
независимо от функции BMP. Транскрипты, кодирующие Foxc1 и Foxc2, которые
ранее было показано, что они необходимы для закрытия век, не обнаруживались
в Smad4 CKO животных. c-Jun, еще один ключевой регулятор
закрытие века, присутствовал и фосфорилировался в клетках перидермы века в
время слияния, но не смог переместиться в ядро в отсутствие BMP
функция. Smad4 CKO мышей также показали преждевременные
дифференцировка эпителия конъюнктивы, гиперплазия конъюнктивы и
приобретение характеристик эпидермиса, в том числе формирование
эктопический ряд волосяных фолликулов на месте мейбомиевых желез. Второй ряд
ресниц является признаком синдрома лимфедема-дистихиаза человека, который
связаны с мутациями в FOXC2.
Транскрипты, кодирующие Foxc1 и Foxc2, которые
ранее было показано, что они необходимы для закрытия век, не обнаруживались
в Smad4 CKO животных. c-Jun, еще один ключевой регулятор
закрытие века, присутствовал и фосфорилировался в клетках перидермы века в
время слияния, но не смог переместиться в ядро в отсутствие BMP
функция. Smad4 CKO мышей также показали преждевременные
дифференцировка эпителия конъюнктивы, гиперплазия конъюнктивы и
приобретение характеристик эпидермиса, в том числе формирование
эктопический ряд волосяных фолликулов на месте мейбомиевых желез. Второй ряд
ресниц является признаком синдрома лимфедема-дистихиаза человека, который
связаны с мутациями в FOXC2.
ВВЕДЕНИЕ
Образование век из поверхностной эктодермы и подлежащей периокулярной области мезенхима включает четыре процесса: спецификацию века, рост, закрытие и повторное открытие.У мышей спецификация век начинается к 9-му дню эмбриона (E), когда экспрессия фактора транскрипции Foxl2 определяет будущее местоположение век дорсально и вентрально по отношению к глобусу (Swindell et al., 2008). В E11.5, инвагинация дорсальной и вентральной периокулярной эктодермы сигнализирует о начало периода роста век. Образовавшиеся складки век разрастаются навстречу друг другу по поверхности глаза между E11.5 и E15.5. В E15.5, выступ наружного перидермального слоя эктодермы расширяется. от краев века через роговицу до соприкосновения расширений перидермы и предохранитель.Два века разделяются при «открытии глаз» примерно на 10-й день после рождения. (P10) (Финдлейтер и др., 1993).
Закрывающиеся веки состоят из слабо организованной мезенхимы и
вышележащий эпителий. Эпителий век дифференцируется на
глазной эпидермис (внешняя поверхность века) и глазной
конъюнктива (внутренняя поверхность века). Глазная конъюнктива
продолжается с бульбарной конъюнктивой (эпителий, покрывающий переднюю
периферии глазного яблока), которая продолжается с эпителием роговицы на
самая передняя поверхность земного шара
(Рис. 1).
1).
После закрытия века эпидермис век дифференцируется как часть кожа. Начинается расслоение и кератинизация, и волосы становятся равномерно расположенными. по краям век образуются фолликулы ресниц. Тем не менее эпителий конъюнктивы расслаивается только после того, как веки снова откроются и остается некератинизированным на протяжении всей жизни. Зрелый эпителий конъюнктивы содержит большое количество бокаловидных клеток, которые производят слизь, важную для свойства слезной пленки.Вскоре после рождения мейбомиевые железы, которые вырабатывают липидный компонент слез, образующихся при росте конъюнктивы эпителиальные клетки около внутренней поверхности края века (Findlater et al., 1993).
Дефекты роста или сращения век могут приводить к открытию век при
рождение (EOB). Требуется удивительное количество генов и сигнальных путей.
для закрытия век. Фенотип EOB наблюдается у мышей с делецией зародышевой линии
активин β-B ( Inhbb ), киназа MEK1 ( Map3k1 ), c-Jun
N-терминальная киназа ( Mapk8 ), c-Jun ( июн ), рост эпидермиса
фактор (EGF) члены семейства HB-EGF ( Hbegf ) и трансформирующий рост
фактор α ( Tgfa ) и их рецептор ( Egfr ), фибробласт
фактор роста 10 ( Fgf10 ), его рецептор ( Fgfr2 ), вилка
факторы транскрипции Foxc1 и Foxc2 , а Wnt
антагонист Dkk2 (Gage et al.,
2008; Кидсон и др.,
1999; Куме и др.,
1998; Ли и др.,
2001; Ли и др.,
2003; Luetteke et al.,
1994; Luetteke et al.,
1993; Miettinen et al.,
1995; Mine et al.,
2005; Смит и др.,
2000; Такатори и др.,
2008; Тао и др.,
2005; Vassalli et al.,
1994; Вестон и др.,
2004; Зенц и др.,
2003; Чжан и др.,
2003 г.). Предыдущие исследования показали, что активин β-B способствует
закрытие века путем активации Smad-независимого каскада с участием MEK-киназы1,
N-концевая киназа Jun (JNK) и c-Jun
(Такатори и др., 2008;
Weston et al., 2004;
Zhang et al., 2003). EGF
члены семьи способствуют миграции перидермы, активируя ERK
сигнальный путь (Mine et al. ,
2005). Перед каскадом EGF c-Jun увеличивает рецептор EGF
экспрессия (Li et al., 2003;
Zenz et al., 2003). FGF10
контролирует пролиферацию эпителия век и миграцию перидермы, стимулируя
экспрессии активина β-B и TGFα, а также путем модуляции
Выражение звукового ежа ( Shh )
(Тао и др., 2005). В
введение антагониста Shh короткого действия на E9 приводит к EOB
(Lipinski et al., 2008).
Недавно мыши, лишенные антагониста Wnt Dkk2 , показали EOB,
указывает на то, что активность Wnt должна быть правильно настроена во время век
разработка.
,
2005). Перед каскадом EGF c-Jun увеличивает рецептор EGF
экспрессия (Li et al., 2003;
Zenz et al., 2003). FGF10
контролирует пролиферацию эпителия век и миграцию перидермы, стимулируя
экспрессии активина β-B и TGFα, а также путем модуляции
Выражение звукового ежа ( Shh )
(Тао и др., 2005). В
введение антагониста Shh короткого действия на E9 приводит к EOB
(Lipinski et al., 2008).
Недавно мыши, лишенные антагониста Wnt Dkk2 , показали EOB,
указывает на то, что активность Wnt должна быть правильно настроена во время век
разработка.
Неясно, передаются ли сигналы костного морфогенетического белка (BMP). играет роль в закрытии век. Фенотип EOB обнаружен у одной мыши. штамм, в котором Bmpr1a был условно удален в эктодерме посредством с использованием трансгена Cre-рекомбиназы, управляемого кератином 14 (Andl et al., 2004). Тем не мение, мыши со сверхэкспрессией антагониста BMP noggin под контролем человека Промоторы K14 или K5 имели дефекты век, но не имели фенотипа EOB (Plikus et al., 2004; Шаров и др., 2003). мышей сверхэкспрессирует ингибирующий Smad Smad7, управляемый бычьим промотором K5, имели фенотип EOB (He et al., 2002). Однако был ли этот фенотип связан с блокирование передачи сигналов TGFβ, активина или BMP не выяснено. В кроме того, сверхэкспрессия антагонистов передачи сигналов BMP или дефицит Сигнальный путь BMP вызывает другие дефекты эпителия, которые могут косвенно результат в EOB. Например, в некоторых из этих случаев эпидермальный, эпителий конъюнктивы и роговицы был гиперпластическим, а потовые железы трансдифференцируется в волосяные фолликулы (Он и др., 2002; Plikus et al., 2004).
Чтобы прояснить возможную функцию передачи сигналов BMP в развитии век, мы
условно удалены два BMP-рецептора I типа, два из BMP-активированных
R-Smads или со-Smad Smad4 в предполагаемом эпителии век, начиная с
E9. В каждом случае у мышей наблюдали нормальное формирование век и адекватный рост.
но эпителий век не слился, что привело к фенотипу EOB.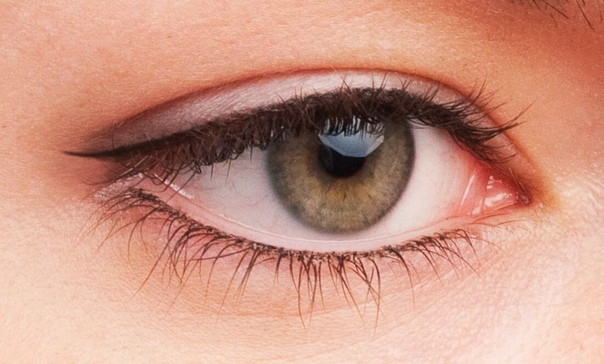 Удаление
единственного рецептора TGFβ типа II или двух активин- и
Активированные TGFβ R-Smads не мешали закрытию век.Дальше
анализ показал, что Fgf10 из мезенхимы активирует Fgfr2 в крышке
эктодерма. Передача сигналов Fgfr2 модулирует уровни Shh, в результате чего получается Bmp4 экспрессия в мезенхиме. Передача сигналов FGF также ингибирует передачу сигналов Wnt в
эктодерма век, независимо от ее влияния на экспрессию BMP. БМП
необходим для экспрессии факторов транскрипции Foxc1 и Foxc2 в
эктодерма, ядерная транслокация активированного c-Jun в клетках перидермы,
правильное время дифференцировки эпителия конъюнктивы и
установление судьбы эпителиальных клеток конъюнктивы.При отсутствии БМП
сигнализируя, эктопические волосяные фолликулы, образующиеся на внутренних краях века в
за счет мейбомиевых желез, особенности человеческого
лимфедема-дистихиазный синдром.
Удаление
единственного рецептора TGFβ типа II или двух активин- и
Активированные TGFβ R-Smads не мешали закрытию век.Дальше
анализ показал, что Fgf10 из мезенхимы активирует Fgfr2 в крышке
эктодерма. Передача сигналов Fgfr2 модулирует уровни Shh, в результате чего получается Bmp4 экспрессия в мезенхиме. Передача сигналов FGF также ингибирует передачу сигналов Wnt в
эктодерма век, независимо от ее влияния на экспрессию BMP. БМП
необходим для экспрессии факторов транскрипции Foxc1 и Foxc2 в
эктодерма, ядерная транслокация активированного c-Jun в клетках перидермы,
правильное время дифференцировки эпителия конъюнктивы и
установление судьбы эпителиальных клеток конъюнктивы.При отсутствии БМП
сигнализируя, эктопические волосяные фолликулы, образующиеся на внутренних краях века в
за счет мейбомиевых желез, особенности человеческого
лимфедема-дистихиазный синдром.
МАТЕРИАЛЫ И МЕТОДЫ
Генотипирование и спаривание мышей
В этом исследовании использовались следующие генетически модифицированные мыши: Le-Cre
(Ashery-Padan et al., 2000), Acvr1 flox (Dudas et al.,
2004), Бмпр1а флокс
(Гауссин и др., 2002), Smad4 flox (Ян и др.,
2002), Smad1 flox
(Хуанг и др., 2002), Smad5 flox (Umans et al.,
2003), Smad2 flox
(Piek et al., 2001), Smad3 нокаут зародышевой линии (Робертс
et al., 2006), Fgfr2 флоксированный
(Yu et al., 2003), Tgfbr2 флоксированный (Chytil et al.,
2002), пресенилин 1 флоксед (Ю.
et al., 2001), нокаут пресенилина 2 зародышевой линии
(Steiner et al., 1999) и
TOPGAL, репортерный штамм Wnt (DasGupta и
Fuchs, 1999).Спаривания мышей, гомозиготных по
floxed аллель, только один из которых был Cre-положительным, привел к пометам у
около половины потомства были Cre-позитивными (условный нокаут,
CKO). Остальные были Cre-отрицательными (дикий тип). Полдень того дня, когда
была обнаружена вагинальная пробка, расценивалась как эмбриональный день (E) 0,5 г. разработка. Эмбрионы были собраны на желаемых стадиях ( n = от 3 до 5
для каждого генотипа и стадии).
разработка. Эмбрионы были собраны на желаемых стадиях ( n = от 3 до 5
для каждого генотипа и стадии).
Гистология
Головки эмбрионов фиксировали в 4% параформальдегиде / PBS в течение ночи при 4 ° C, обезвоженный через серию концентраций этанола, залитый в парафин и срезы толщиной 4 мкм.Для морфологических исследований разрезы были окрашены гематоксилином и эозином (Surgipath, Richmond, IL, USA).
Гибридизация in situ
Замороженные срезы фиксировали в 4% параформальдегиде / PBS, обрабатывали протеиназа K (10 мкг / мл), постфиксированная в 4% параформальдегиде / PBS и ацетилированная в растворе триэтаноламин-уксусного ангидрида. Образцы были предварительно гибридизированы в 50% формамид, 5 × SSC, 5 мМ EDTA, 1 × Denhardt’s, 100 мкг / мл гепарина, 0,3 мг / мл дрожжевой тРНК и 0.1% Твин-20, инкубированный в том же растворе с рибозонды в течение ночи, промывают 0,2 × SSC, блокируют 10% сывороткой ягненка и инкубировали с антителом к дигоксигенину в течение ночи. Цветовая реакция была разработан с использованием NBT и BCIP в темноте. После завершения реакции предметные стекла промывали PBS, фиксировали в 4% параформальдегиде / PBS и помещали в 100% глицерин.
меченных дигоксигенином рибозондов были синтезированы из кДНК, полученной из РНК, выделенная из век E15.5 дикого типа с использованием следующего праймера для ПЦР пар:
Foxc1, 5′-CCAGAAAGTGTTCCAAAAGC-3’и 5′-GAAACCACCCCAGACTAATG-3 ‘;
Foxc2, 5′-GCCACCTCCTGGTATCTGAAC-3 ‘и 5’-CTGGGCAAGAACAAAATAGCC-3 ‘;
BMP4, 5′-TGGTAACCGAATGCTGATGG-3 ‘и 5’-GGCGACGGCAGTTCTTATTC-3 ‘;
Sfrp1, 5′-ATCCCCCTCTTTCTGCCTTAG-3 ‘и 5’-GAAATACCTCTGGGCACTTGG-3 ‘;
Dkk2, 5′-TTTACAAAGTGGGTTCCCTTG-3 ‘и 5’-CTCCATTTTCACATCACAAAGC-3 ‘.

Зонд для исправления 1 был любезным подарком доктора Дэвида Орница (Вашингтон). Университет, Сент-Луис, Миссури, США). Паттерны экспрессии генов сравнивали между CKO и однопометники дикого типа и каждую гибридизацию in situ проводили в минимум в два раза.
Иммунофлуоресцентное окрашивание
Замороженные срезы нагревали до комнатной температуры, а затем фиксировали в 4% параформальдегид / PBS. После трех промывок в PBS образцы обрабатывали 3% H 2 O 2 в метаноле для тушения эндогенной пероксидазы активность, заблокированная в 5% козьей сыворотке / 0.1% Triton X-100, инкубированный в первичном антитело в течение ночи, промывали и обрабатывали амплификацией тирамида. В антитела к pSmad1 / 5/8 и p-c-Jun были получены от Cell Signaling Technology. (Дэнверс, Массачусетс, США). Антитела кератина 14 и кератина 10 были получены от Covance. Research Products (Денвер, Пенсильвания, США). Антитело кератина 4 было получено из Сигма-Олдрич (Сент-Луис, Миссури, США). Антитело Dkk2 было из Санта-Крус. Биотехнология (Санта-Крус, Калифорния, США).
Окрашивание X-gal
Стадийные эмбрионы, экспрессирующие репортерный ген lacZ , были зафиксированы на 4% в параформальдегид / PBS при 4 ° C в течение 30 минут, дважды промывали в PBS с 2 мМ MgCl 2 ,0.02% NP-40 / 0,01% дезоксихолата (DOC) и окрашено X-gal раствор [5 мМ K3Fe (CN) 6 , 5 мМ K4Fe (CN) 6 , 1 M MgCl 2 , 0,02% NP-40 / 0,01% DOC NP-40, 1 мг / мл X-гал в PBS] для 5 часов при 37 ° C, после фиксации 4% параформальдегидом в течение 1 часа, криозащищенные и, при необходимости, готовили срезы размером 10 мкм.
Окрашивание и количественное определение BrdU и TUNEL
Беременным самкам мышей вводили 50 мг / кг смеси 10 мМ BrdU.
(Roche, Indianapolis, IN, USA) и 1 мМ 5-фтор-5′-дезоксиуридина (Sigma,
Сент-Луис, Миссури, США) и приносили в жертву через 1 час. Моноклональный анти-BrdU
антитело (разведенное 1: 250; Dako, Carpinteria, CA, USA) использовали с
Набор Vectastain Elite Mouse IgG ABC. Срезы контрастировали с
Гематоксилин. Дезоксиуридин, опосредованный терминальной дезоксинуклеотидилтрансферазой (TdT)
Мечение концов трифосфатных никеля (TUNEL) проводили с помощью набора Apoptag (Chemicon,
Темекула, Калифорния, США). Депарафинизированные слайды обрабатывали 3%
H 2 O 2 в метаноле в течение 30 минут, затем протеиназа
К (20 мкг / мл) в течение 15 минут.Слайды инкубировали с TdT.
фермент в уравновешивающем буфере в течение 1 часа при 37 ° C. Реакция была
прекращено промывочным буфером, предоставленным производителем, в течение 10 минут при
комнатная температура. Конъюгат анти-дигоксигенин-пероксидаза добавляли на 30 минут.
минут при комнатной температуре, затем DAB и H 2 O 2 лечение. Слайды контрастировали гематоксилином.
Моноклональный анти-BrdU
антитело (разведенное 1: 250; Dako, Carpinteria, CA, USA) использовали с
Набор Vectastain Elite Mouse IgG ABC. Срезы контрастировали с
Гематоксилин. Дезоксиуридин, опосредованный терминальной дезоксинуклеотидилтрансферазой (TdT)
Мечение концов трифосфатных никеля (TUNEL) проводили с помощью набора Apoptag (Chemicon,
Темекула, Калифорния, США). Депарафинизированные слайды обрабатывали 3%
H 2 O 2 в метаноле в течение 30 минут, затем протеиназа
К (20 мкг / мл) в течение 15 минут.Слайды инкубировали с TdT.
фермент в уравновешивающем буфере в течение 1 часа при 37 ° C. Реакция была
прекращено промывочным буфером, предоставленным производителем, в течение 10 минут при
комнатная температура. Конъюгат анти-дигоксигенин-пероксидаза добавляли на 30 минут.
минут при комнатной температуре, затем DAB и H 2 O 2 лечение. Слайды контрастировали гематоксилином.
Анатомия века мыши дикого типа на E15.5. Компоненты нормальный глазной эпителий имеет цветовую маркировку: бульбарная конъюнктива — пурпурный; пальпебральный конъюнктива розовая; эпидермис век, темно-красный; перидерма, апельсин; роговица синий.
В поверхностном эпителии глаза подсчитывалиBrdU и TUNEL-положительных клеток. на E14.5 в 4-5 срезах от каждого эмбриона (дикий тип, n = 6; Smad4 CKO , n = 8). Средние и стандартная ошибка (s.e.m.) были рассчитывается на основе объединенных данных. Различия считались значимыми, если П <0.05, по результатам теста Стьюдента t .
РЕЗУЛЬТАТЫ
Передача сигналов BMP необходима для закрытия века мыши
Для изучения функций BMP в раннем развитии глаз мы использовали Cre
рекомбиназа для удаления floxed аллелей ключевых компонентов передачи сигналов BMP
путь. Трансгенная экспрессия Cre управлялась промотором, который является первым
экспрессируется на E9 в плакоде хрусталика и в эктодерме, которая позже
дифференцируется в эпителий глазной поверхности (Le-Cre)
(Ашери-Падан и др., 2000).
Эпителий глазной поверхности, на который нацелен трансген, включает пальпебральный
эпидермис, конъюнктива век, конъюнктива глазного яблока и эпителий роговицы
(Ашери-Падан и др., 2000)
(Рисунок 1). Мыши Le-Cre были
скрещивается с мышами с floxed аллелями двух из трех рецепторов BMP типа I.
( Acvr1 и Bmpr1a ), два активированных BMP R-Smads
( Smad1 и Smad5 ), два активина / TGFβ-активированных
R-Smads ( Smad2 и Smad3 ), единственный тип II TGFβ
рецептор ( Tgfbr2 ) или со-Smad Smad4 .Спаривания были между
Cre-положительные гомозиготные животные флокса и Cre-отрицательные гомозиготные животные flox,
уверяя, что около половины потомства были Cre-положительными и не имели животных
получили две копии трансгена.
Трансгенная экспрессия Cre управлялась промотором, который является первым
экспрессируется на E9 в плакоде хрусталика и в эктодерме, которая позже
дифференцируется в эпителий глазной поверхности (Le-Cre)
(Ашери-Падан и др., 2000).
Эпителий глазной поверхности, на который нацелен трансген, включает пальпебральный
эпидермис, конъюнктива век, конъюнктива глазного яблока и эпителий роговицы
(Ашери-Падан и др., 2000)
(Рисунок 1). Мыши Le-Cre были
скрещивается с мышами с floxed аллелями двух из трех рецепторов BMP типа I.
( Acvr1 и Bmpr1a ), два активированных BMP R-Smads
( Smad1 и Smad5 ), два активина / TGFβ-активированных
R-Smads ( Smad2 и Smad3 ), единственный тип II TGFβ
рецептор ( Tgfbr2 ) или со-Smad Smad4 .Спаривания были между
Cre-положительные гомозиготные животные флокса и Cre-отрицательные гомозиготные животные flox,
уверяя, что около половины потомства были Cre-положительными и не имели животных
получили две копии трансгена.
В каждом нокауте, нацеленном на путь BMP ( Acvr1 / Bmpr1a, Smad1 / 5 и Smad4 ), Cre-позитивные животные имели открытое веко при рождении (EOB)
фенотип (рис. 2B-D).
Потомство с условной делецией одного аллеля генов пути BMP
( Acvr1 / Bmpr1a, Smad1 / 5 и Smad4 ; не показаны), оба Аллели Tgfbr2 (рис.2G) или оба аллеля Smad2 и Smad3 (данные не
показаны) при рождении имели нормальный вид, закрытые веки и нормальный вид
эпителий конъюнктивы между E15.5 и рождением. Изучая эмбрионы между
E16.5 и рождения, мы обнаружили, что веки Cre-позитивных эмбрионов с EOB
никогда не закрывался, что указывает на то, что фенотип возник в результате неспособности
закрытие века, а не из-за преждевременного открытия века (не показано). Потому что только
нокауты в каноническом пути BMP-Smad имели фенотип EOB, в
в остальных описанных исследованиях мы показываем только фенотип Smad4 CKO мышей для представления функции передачи сигналов BMP
при закрытии век. Все показанные «контрольные» веки принадлежат гомозиготному флоксу,
Cre-отрицательные однопометники.
Все показанные «контрольные» веки принадлежат гомозиготному флоксу,
Cre-отрицательные однопометники.
Дефекты век у мышей с дефицитом передачи сигналов BMP. ( A-G ) Окрашивание гематоксилином и эозином фронтальных срезов глаза из дикого типа (A, E), Acvr1; Bmpr1a DCKO (B), Smad4 CKO (C, F), Smad1 / 5 DCKO (D) и Tgfbr2 CKO (G) мышей в постнатальный день 3 (P3).Дикого типа веки слиты (стрелка в A), тогда как веки дефицитны по передаче сигналов BMP в эктодерма отдельные (стрелки на B-D). (E, F) Более высокое увеличение виды вставок в A и C. (E) Кератин, окрашенный эозином (стрелка) и Кератиноциты, окрашенные гематоксилином (наконечник стрелки) в веко век дикого типа эпидермис. (F) Эктопический кератиноподобный белок, окрашенный эозином (стрелка) и эктопический Окрашенные гематоксилином кератиноцитоподобные клетки (стрелка) в Smad4 CKO конъюнктива. Звездочки на E и F обозначают гиперплазия конъюнктивы Smad4 CKO .(ГРАММ) Tgfbr2 CKO Веки новорожденного срослись (стрелка). Масштабные полосы: 100 мкм.
Помимо фенотипа EOB, глаза с недостаточностью передачи сигналов BMP показали гиперплазия и ороговение конъюнктивы эпителий. В отличие от двухслойного эпителия, наблюдаемого у животных дикого типа эмбрионов эпителий конъюнктивы состоял из нескольких слоев клеток, в том числе гранулированный слой с темными, окрашенными гематоксилином гранулами кератогиалина, и эозинофильный роговой слой, свидетельствующий о кератинизации (Рис.2E, F).
Рис. 3. Клеточная пролиферация в Smad4 CKO эпителий век в
E14.5. ( A-D ) Окрашивание BrdU в лобных срезах глаза дикого типа
верхнее веко (A), нижнее веко дикого типа (C), Smad4 CKO верхнее веко (B) и Smad4 CKO нижнее веко (D). Красные линии
обозначить примерные границы бульбарной и конъюнктивы век и
эпидермис век.( E ) Процент клеток, меченных BrdU, в каждой
отсек. По сравнению с животными дикого типа индекс маркировки BrdU в Smad4 CKO бульбарный и глазной эпителий конъюнктивы
значительно увеличился. Оси и указывают средний процент
Включение BrdU в каждой исследуемой области. Планки погрешностей представляют собой среднюю погрешность. * P <0,05, *** P <0,001. Б, бульбарная
конъюнктива; ПК, конъюнктива глазного дна; ПЭ, глазной эпидермис.Шкала шкалы в
A: 100 мкм.
Красные линии
обозначить примерные границы бульбарной и конъюнктивы век и
эпидермис век.( E ) Процент клеток, меченных BrdU, в каждой
отсек. По сравнению с животными дикого типа индекс маркировки BrdU в Smad4 CKO бульбарный и глазной эпителий конъюнктивы
значительно увеличился. Оси и указывают средний процент
Включение BrdU в каждой исследуемой области. Планки погрешностей представляют собой среднюю погрешность. * P <0,05, *** P <0,001. Б, бульбарная
конъюнктива; ПК, конъюнктива глазного дна; ПЭ, глазной эпидермис.Шкала шкалы в
A: 100 мкм.
EOB, вызванный прерыванием передачи сигналов BMP, не связан с уменьшением пролиферация клеток или повышенная гибель клеток
Мы исследовали, способствовала ли потеря передачи сигналов BMP отказу закрытие век, влияя на пролиферацию или гибель клеток. На E12.5, BrdU и маркировка TUNEL была аналогична для дикого типа и Smad4 CKO веки (не показаны). Однако произошло значительное увеличение маркировки BrdU. в конъюнктивальном эпителии Smad4 CKO глаз на E14.5 (Рис. 3A-E). Это наблюдение соответствовало гиперплазии конъюнктивы, наблюдаемой на более поздних стадиях. Нет увеличение мечения BrdU было обнаружено в эпидермисе глазного дна (Рис. 3A-E), и никаких изменений не было. наблюдается в запрограммированной гибели клеток в любом поверхностном эпителии глаза (не показано).
передача сигналов BMP активирует экспрессию факторов транскрипции, которые необходимы для закрытия век
Экспрессия факторов транскрипции вилки Foxc1 и Foxc2
требуется для закрытия век (Kidson et al.
al., 1999; Куме и др.,
1998; Смит и др.,
2000). Foxc1 мРНК присутствовала в верхних и нижних клетках дикого типа.
Эпителий нижнего века (рис.
4A), но не обнаруживается в Smad4 CKO веках
(Рис. 4C). Точно так же в
эмбрионы дикого типа, мРНК Foxc2 экспрессировалась в пальпебральной конъюнктиве
эпителиальные клетки (рис. 4D), но
не может быть обнаружен в Smad4 CKO конъюнктиве глазного яблока
(Рис. 4F).
4C). Точно так же в
эмбрионы дикого типа, мРНК Foxc2 экспрессировалась в пальпебральной конъюнктиве
эпителиальные клетки (рис. 4D), но
не может быть обнаружен в Smad4 CKO конъюнктиве глазного яблока
(Рис. 4F).
передача сигналов BMP необходима для обеспечения транслокации c-Jun в ядра мигрирующих клеток перидермы
c-Jun и сигнальный каскад, который приводит к его фосфорилированию: требуется для закрытия век (Li et al., 2003; Зенц и др., 2003 г.). У мышей дикого типа полка клеток перидермы начинает расти. выходят за пределы век на E15.0 (Рис. 5A), покрывая большую часть роговица на E15.5 (рис. 5B). В ядра этих клеток перидермы были сильно окрашены антителами к фосфорилированный c-Jun (рис. 5А, Б, врезки). В веко Smad4 CKO эпителий, внешний вид края век был сравним с таковым у дикого тип на E15.0 (рис. 5C), хотя на ст. E15.5 мигрирующих клеток перидермы было меньше, чем в контрольные глаза (рис.5D). в Smad4 CKO эмбрионов, уровни фосфорилированного c-Jun оказался ниже, чем в клетках перидермы дикого типа, и окрашивание p-c-Jun было присутствует в перинуклеарной цитоплазме, но не в ядрах (Рис. 5В, Г, врезки). Таким образом, БМП сигнализация необходима для полной активации c-Jun и для его транслокация в ядро для выполнения своей функции транскрипции фактор.
Экспрессия и функция BMP регулируются передачей сигналов FGF во время закрытие века
Передача сигналов Fgf10 через Fgfr2 необходима для роста и закрытия век.
(Ли и др., 2001;
Тао и др., 2005). К
определить, существует ли связь между передачей сигналов FGF и BMP, мы
удалил Fgfr2 в предполагаемом эпителии века с помощью Le-Cre. мышей
дефицит Fgfr2 в эктодерме показал фенотип EOB, как
описано ранее (Garcia et al.,
2005), а также недостаточность экспрессии и функции BMP. В
нижних век дикого типа на ст. E15.5, мРНК Bmp4 экспрессировалась в
скопление мезенхимальных клеток, лежащих в основе эпителия конъюнктивы глазного яблока
(Рис. 6А, стрелки), но Bmp4 транскриптов не обнаруживались в нижних веках Fgfr2 CKO мышей (рис.
6С). Bmp4 мРНК экспрессировалась в двух группах мезенхимальных
клетки верхнего века дикого типа: в кластере, соответствующем найденным
в нижнем веке (рис. 6А,
стрелки) и кластером под эпидермисом глазного дна
(Рис. 6A, стрелки). в Fgfr2 CKO верхнее веко, Bmp4 накопление мРНК в
мезенхима, лежащая под конъюнктивой век, не была затронута, но Bmp4 транскриптов не были обнаружены в мезенхиме, лежащей в основе
эпидермис век (рис.6Б).
6А, стрелки), но Bmp4 транскриптов не обнаруживались в нижних веках Fgfr2 CKO мышей (рис.
6С). Bmp4 мРНК экспрессировалась в двух группах мезенхимальных
клетки верхнего века дикого типа: в кластере, соответствующем найденным
в нижнем веке (рис. 6А,
стрелки) и кластером под эпидермисом глазного дна
(Рис. 6A, стрелки). в Fgfr2 CKO верхнее веко, Bmp4 накопление мРНК в
мезенхима, лежащая под конъюнктивой век, не была затронута, но Bmp4 транскриптов не были обнаружены в мезенхиме, лежащей в основе
эпидермис век (рис.6Б).
Предыдущее исследование показало, что Fgf10 поддерживает экспрессию Shh в мезенхима века (Tao et al., 2005). Мы обнаружили, что еж функционирует в мезенхиме, так как измеряется по экспрессии патченного рецептора hedgehog 1 ( Ptch2 ), прямая цель передачи сигналов Shh, была удивительно похожа на распределение Bmp4 транскриптов (Рис. 6D). Более того, паттерн остаточной экспрессии Ptch2 в Fgfr2 CKO веко было похоже на рисунок остаточного Bmp4 выражение.Хотя экспрессия Ptch2 сохранилась в эктодерма и мезенхима верхнего века, в нижнем веке она значительно уменьшилась эктодерма и мезенхима Fgfr2 CKO мышей (Рис. 6E). ПТЧ2 экспрессия не была затронута у мышей Smad4 CKO (Рис. 6F).
В соответствии с зависимостью экспрессии Bmp4 от Fgfr2,
ядерное окрашивание фосфорилированных Smad1 / 5/8, рецептор-активируемых Smads
которые передают сигналы BMP, были сильными в эпителиальных клетках дикого типа
(Рис.6G, H), но сильно
уменьшено в верхнем и нижнем веке у мышей Fgfr2 CKO (Рис. 6I, J). Как в Smad4 CKO век, Foxc1 и Foxc2 мРНК
не обнаружен в Fgfr2 CKO эпителии конъюнктивы
(Рис. 4B, E). Таким образом, FGF
сигнализация управляет закрытием век, по крайней мере частично, посредством активации
Сигнализация BMP.
Foxc1 и Foxc2 транскрипты значительно сокращены в Fgfr2 CKO и Smad4 CKO века. ( A-C ) Гибридизация in situ для мРНК Foxc1 на лобном глазу секции дикого типа (A), Fgfr2 CKO (B) и Smad4 CKO (C) веки на E15.5. В век дикого типа (A), Foxc1 мРНК была обнаружена в глазном эпителии, пальпебральном конъюнктива, бульбарная конъюнктива, перидерма и сетчатка. В Fgfr2 CKO (B) и Smad4 CKO (C) веки, Foxc1 мРНК обнаруживалась только в сетчатке.( D-F ) На месте гибридизация мРНК Foxc2 на лобных срезах глаза дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) веки. В веках дикого типа (D) мРНК Foxc2 была обнаружена в конъюнктива век и сетчатка на E15.5. В Fgfr2 CKO (E) и Smad4 CKO (F) век, Foxc2 мРНК была обнаруживается только в сетчатке. Масштабные линейки: 100 мкм.
Активация Fgfr2 подавляет передачу сигналов Wnt и способствует экспрессия
Sfrp1 BMP-независимым образом Потеря Dkk2, антагониста передачи сигналов Wnt, вызывает EOB, показывая, что
чрезмерная передача сигналов Wnt может предотвратить закрытие век
(Гейдж и др., 2008). К
далее оценить регуляцию передачи сигналов пути Wnt на поверхности глаза
epithelia, мы произвели условные нокауты Fgfr2 и Smad4 в эктодерме на фоне TOPGAL, в котором каноническая передача сигналов Wnt
активирует трансген-репортер β-галактозидазы
(ДасГупта и Фукс, 1999).В
веки дикого типа (Cre-отрицательные) на ст. E15.5, окрашивание β-галактозидазой было
в изобилии в волосяных фолликулах эпидермиса и в эпителии конъюнктивы
полосой у края верхнего века. Более слабое окрашивание присутствовало в
полоса по краю нижнего века
(Рис. 7A, D). В Fgfr2 CKO глаза, активность репортера TOPGAL увеличилась в
интенсивность и распространение по эпителию конъюнктивы верхнего и нижнего
веки (рис. 7Б, Д). В Smad4 CKO веки, экспрессии β-галактозидазы не было
увеличилось, но имело другое распространение от дикого типа.Вместо
локализующиеся сплошной полосой в периферическом эпителии конъюнктивы,
окрашивание присутствовало в дополнительном ряду эктопических волосяных фолликулов в верхнем
и нижние веки (рис. 7C, F).
Двойной ряд ресниц называется дистихиазом. Поскольку возникает дистихиаз
у человека и мышей гаплонедостаточно для FOXC2
(Fang et al., 2000;
Kriederman et al., 2003), это
открытие согласуется с нашим наблюдением, что передача сигналов BMP необходима для Foxc2 выражение.
Более слабое окрашивание присутствовало в
полоса по краю нижнего века
(Рис. 7A, D). В Fgfr2 CKO глаза, активность репортера TOPGAL увеличилась в
интенсивность и распространение по эпителию конъюнктивы верхнего и нижнего
веки (рис. 7Б, Д). В Smad4 CKO веки, экспрессии β-галактозидазы не было
увеличилось, но имело другое распространение от дикого типа.Вместо
локализующиеся сплошной полосой в периферическом эпителии конъюнктивы,
окрашивание присутствовало в дополнительном ряду эктопических волосяных фолликулов в верхнем
и нижние веки (рис. 7C, F).
Двойной ряд ресниц называется дистихиазом. Поскольку возникает дистихиаз
у человека и мышей гаплонедостаточно для FOXC2
(Fang et al., 2000;
Kriederman et al., 2003), это
открытие согласуется с нашим наблюдением, что передача сигналов BMP необходима для Foxc2 выражение.
Хотя потеря экспрессии Dkk2 вызывает EOB
(Гейдж и др., 2008), Dkk2 мРНК или экспрессия белка не были затронуты в Fgfr2 CKO или Smad4 CKO веко
эпителиальные клетки (рис. 8A-F).
Передача сигналов FGF модулирует экспрессию Shh в веках и в других
тканях, передача сигналов hedgehog индуцирует экспрессию антагониста Wnt,
секретируемый белок 1, связанный с фризлом (Sfrp1)
(He et al., 2006;
Като и Като, 2006). Мы
исследовали уровни мРНК Sfrp1 в организме дикого типа, Fgfr2 CKO и Smad4 CKO века.В
веки дикого типа, мРНК Sfrp1 экспрессировалась во всем веке
эпителий, с наибольшей экспрессией в эпителии конъюнктивы
(Рис. 8G). В соответствии с
эффекты передачи сигналов FGF и BMP на активность TOPGAL, Sfrp1 транскрипты не обнаруживались в Fgfr2 CKO веках
(Рис. 8H), но присутствовали в
нормальные уровни в Smad4 CKO веках
(Рис. 8I). Таким образом, передача сигналов FGF
подавляет передачу сигналов Wnt, по крайней мере частично, посредством активации Сфрп1 . Однако контроль экспрессии Sfrp1 с помощью Fgfr2 является
независимо от передачи сигналов BMP.
Однако контроль экспрессии Sfrp1 с помощью Fgfr2 является
независимо от передачи сигналов BMP.
Ядерная локализация фосфорилированного c-Jun требует Smad4 . Иммуноокрашивание фосфорилированного c-Jun (p-c-Jun, зеленый) на лобных срезах глаза от E15.0 (A) дикого типа, E15.5 (B) дикого типа, Smad4 CKO E15.0 (C) и Smad4 CKO E15.5 (D) эмбрионы.( A ) В веках дикого типа небольшое количество мигрирующих клетки перидермы присутствуют на E15.0. Окрашивание на p-c-Jun присутствует в ядра этих клеток (вставка). ( B ) По E15.5 количество перидермы клеток увеличилось, и они начали мигрировать по роговице. Окрашивание поскольку p-c-Jun присутствует в ядрах этих клеток (вставка). ( C ) При E15.0, клетки перидермы Smad4 CKO век оказались похожими по количеству к дикому типу. Однако окрашивание p-c-Jun было ограничено цитоплазма вокруг ядер (вставка).( D ) На ст. E15.5 меньше клеток перидермы в веках Smad4 CKO присутствовали , чем в веках дикого типа. Окрашивание на p-c-Jun было слабее, чем на клетки перидермы дикого типа, и все еще оставалось ограничивается перинуклеарной цитоплазмой (вставка). На вставках показано окрашивание p-c-Jun. без ядерного контрастного красителя (синий). Шкала в A: 100 мкм.
Хотя путь BMP-Smad не участвует в подавлении передачи сигналов Wnt,
по-видимому, он необходим для подавления образования эктопических волосяных фолликулов из
эпителиальные клетки конъюнктивы по внутреннему краю века
(Рис.7C). В его отсутствие
равномерная полоса передачи сигналов Wnt у края века была заменена рядом
ресничные фолликулы (рис. 7C, F).
Эти наблюдения предполагают, что передача сигналов BMP поддерживает паттерн Wnt
активность пути, необходимая для дифференцировки мейбомиевой железы
клетки-предшественники на внутренних краях век.
Передача сигналов BMP подавляет дифференцировку конъюнктивального эпителиальные клетки до закрытия век и определяет эпителиальный конъюнктивальный клеточная судьба
Эпителий конъюнктивы у мышей с дефицитом передачи сигналов BMP развился. черты, напоминающие эпидермис, в том числе ороговение и эктопические волосяные фолликулы.Поэтому мы исследовали уровни эпителиального маркер дифференцировки клеток кератин 14 (K14) на ст. E15.0, до закрытия века, и на E15.5, во время закрытия. Мы также окрашивали конкретный эпидермис. маркер дифференцировки, кератин 10 (K10) и конъюнктивальный эпителиальный маркер дифференцировки, кератин 4 (K4), на ст. E17.5, после закрытия века. В у мышей дикого типа эпителий конъюнктивы сначала экспрессировал K14 примерно на время закрытия века (рис. 9А). Однако в эпителиальных клетках конъюнктивы с дефицитом BMP передача сигналов, экспрессия K14 была преждевременной (Рис.9C), с мутантом конъюнктива дифференцируется одновременно с эпидермисом. В диком типе E17.5 глаза, K10 экспрессируется эпидермисом, а K4 выражается конъюнктивой. (Рис. 9D, G). Однако в Smad4 CKO эпителий конъюнктивы, мы обнаружили эктопический K10 экспрессия, при этом окрашивание K4 обнаруживается только в нескольких остаточных клетках (Рис. 9F, I), предполагая, что большинство мутантных клеток конъюнктивы трансдифференцировались в клетки эпидермиса. Хотя дифференцировка конъюнктивы в Fgfr2 CKO глазах была преждевременные, что определяется выражением K14 (Рис.9Б), трансдифференцировка конъюнктивы в эпидермис не была очевидной, поскольку Fgfr2 CKO конъюнктива не экспрессирует K10 (Рис. 9E, H). Кажется возможным что остаточная передача сигналов BMP в конъюнктиве Fgfr2 CKO сохранял судьбу клеток конъюнктивы, но был неспособен подавить преждевременные дифференциация этой ткани.
Рис. 6. Передача сигналов от Fgfr2 способствует экспрессии Bmp4, patched 1 и BMP
сигнализация в веко. ( A-C ) Гибридизация in situ для Bmp4 транскриптов на E15.5 в лобных срезах глаза дикого типа
верхнее и нижнее веко (A) и Fgfr2 CKO верхнее (B) и
нижние (C) веки. (A) В веках дикого типа Bmp4 транскриптов
присутствовали в скоплении клеток мезенхимы, лежащих в основе глазного яблока.
конъюнктива (стрелки) и эпидермис век (наконечник стрелки). (B) В верхней
веки Fgfr2 CKO мышей, Bmp4 транскриптов были
обнаруживается в мезенхиме, лежащей под конъюнктивой глазного яблока (стрелка), но
не в мезенхиме, лежащей в основе эпидермиса глазного дна (наконечник стрелки).(C) В
нижнее веко мышей Fgfr2 CKO , Bmp4 транскрипты не были обнаружены в мезенхиме, лежащей в основе пальпебрального
конъюнктива (стрелка). ( D-F ) Гибридизация in situ для Ptch2 транскрипты в лобных срезах глаза дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) века.
У дикого типа (D) и Smad4 CKO век (F), Ptch2 транскрипты присутствовали в кластерах клеток мезенхимы и вышележащих
конъюнктива век (стрелки) и эпидермис век (наконечник стрелки).(E) В
верхние веки Fgfr2 CKO мышей, Ptch2 транскриптов
были уменьшены в мезенхиме и вышележащей конъюнктиве глазного яблока
(стрелка) и едва обнаруживались в мезенхиме и вышележащих
глазной эпидермис (наконечник стрелки). В нижних веках Fgfr2 CKO мышей, Ptch2 транскриптов были
не обнаруживается в мезенхиме и вышележащей конъюнктиве глазного яблока
(стрелка). ( G-J ) Иммуноокрашивание для фосфорилированного Smad1 / 5/8 (pSmad1 / 5/8)
в лобных отделах глаза от верхнего (G) дикого типа, нижнего (H) дикого типа, Fgfr2 CKO верхний (I) и Fgfr2 CKO нижний
(J) веки.(G) В верхнем веке дикого типа, ядерное окрашивание pSmad1 / 5/8
был замечен в мезенхиме века, на конъюнктиве верхнего века
(стрелка), глазной эпидермис (стрелка) и перидерма (звездочка).
( A-C ) Гибридизация in situ для Bmp4 транскриптов на E15.5 в лобных срезах глаза дикого типа
верхнее и нижнее веко (A) и Fgfr2 CKO верхнее (B) и
нижние (C) веки. (A) В веках дикого типа Bmp4 транскриптов
присутствовали в скоплении клеток мезенхимы, лежащих в основе глазного яблока.
конъюнктива (стрелки) и эпидермис век (наконечник стрелки). (B) В верхней
веки Fgfr2 CKO мышей, Bmp4 транскриптов были
обнаруживается в мезенхиме, лежащей под конъюнктивой глазного яблока (стрелка), но
не в мезенхиме, лежащей в основе эпидермиса глазного дна (наконечник стрелки).(C) В
нижнее веко мышей Fgfr2 CKO , Bmp4 транскрипты не были обнаружены в мезенхиме, лежащей в основе пальпебрального
конъюнктива (стрелка). ( D-F ) Гибридизация in situ для Ptch2 транскрипты в лобных срезах глаза дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) века.
У дикого типа (D) и Smad4 CKO век (F), Ptch2 транскрипты присутствовали в кластерах клеток мезенхимы и вышележащих
конъюнктива век (стрелки) и эпидермис век (наконечник стрелки).(E) В
верхние веки Fgfr2 CKO мышей, Ptch2 транскриптов
были уменьшены в мезенхиме и вышележащей конъюнктиве глазного яблока
(стрелка) и едва обнаруживались в мезенхиме и вышележащих
глазной эпидермис (наконечник стрелки). В нижних веках Fgfr2 CKO мышей, Ptch2 транскриптов были
не обнаруживается в мезенхиме и вышележащей конъюнктиве глазного яблока
(стрелка). ( G-J ) Иммуноокрашивание для фосфорилированного Smad1 / 5/8 (pSmad1 / 5/8)
в лобных отделах глаза от верхнего (G) дикого типа, нижнего (H) дикого типа, Fgfr2 CKO верхний (I) и Fgfr2 CKO нижний
(J) веки.(G) В верхнем веке дикого типа, ядерное окрашивание pSmad1 / 5/8
был замечен в мезенхиме века, на конъюнктиве верхнего века
(стрелка), глазной эпидермис (стрелка) и перидерма (звездочка). (ЧАС)
В нижнем веке дикого типа окрашивание pSmad1 / 5/8 наблюдалось на веке
мезенхима, вышележащая конъюнктива глазного дна (стрелка) и перидерма
(звездочка). (I) В верхнем веке эмбрионов Fgfr2 CKO ,
Окрашивание pSmad1 / 5/8 уменьшилось в конъюнктиве глазного яблока (стрелка) и
сильно уменьшен в эпидермисе глазного дна (наконечник стрелки).(J) В Fgfr2 CKO нижнее веко, окрашивание pSmad1 / 5/8 было сильным
снижается в конъюнктиве глазного яблока (стрелка). ul, верхнее веко; ll, нижний
веко. Масштабные линейки: 100 мкм.
(ЧАС)
В нижнем веке дикого типа окрашивание pSmad1 / 5/8 наблюдалось на веке
мезенхима, вышележащая конъюнктива глазного дна (стрелка) и перидерма
(звездочка). (I) В верхнем веке эмбрионов Fgfr2 CKO ,
Окрашивание pSmad1 / 5/8 уменьшилось в конъюнктиве глазного яблока (стрелка) и
сильно уменьшен в эпидермисе глазного дна (наконечник стрелки).(J) В Fgfr2 CKO нижнее веко, окрашивание pSmad1 / 5/8 было сильным
снижается в конъюнктиве глазного яблока (стрелка). ul, верхнее веко; ll, нижний
веко. Масштабные линейки: 100 мкм.
ОБСУЖДЕНИЕ
Smad-зависимая передача сигналов BMP необходима для закрытия века
Условная делеция генов, кодирующих два из трех BMP I типа рецепторов, два из трех BMP-ассоциированных R-Smad или co-Smad Smad4 в эпителий век привел к фенотипу EOB, преждевременному развитию конъюнктивы эпителиальная дифференцировка и трансдифференцировка конъюнктивы в эпидермис, включая формирование фолликулов эктопических ресниц и специфичные для эпидермиса кератиновое выражение.Хотя передача сигналов активина важна для век закрытия, предыдущее исследование показало, что сигналы активина β-B являются трансдуцируется в c-Jun с помощью MEKK1 и JNK, а не через канонический путь Smad (Zhang et al., 2003). Наш результаты согласуются с этой интерпретацией, поскольку удаление R-Smads Smad2 и Smad3, стимулированные активином / TGFβ, не вызывали EOB фенотип. Делеция рецептора TGFβ типа II также не показала фенотип века, что указывает на то, что фенотип Smad4 был обусловлен дефектами в передача сигналов через канонический путь BMP, а не через другой Пути суперсемейства TGFβ, которые разделяют функцию Smad4.
Фенотип EOB век, лишенный эпителиальных рецепторов BMP и
BMP-активированный R-Smads соответствует результатам Bmpr1a делеция с использованием трансгена K14-Cre (Andl et al.
al., 2004) и сверхэкспрессия ингибирующего Smad Smad7
с использованием бычьего промотора K5 (He et al. ,
2002). Неспособность вызванной K14 сверхэкспрессии noggin вызывать
EOB может быть вызван поздней экспрессией K14 в конъюнктиве, так же как
веко закрывается, не давая мозгу времени для предотвращения
активация рецепторов BMP.EOB у мышей с дефицитом передачи сигналов BMP был
не из-за снижения пролиферации клеток или увеличения гибели клеток в
глазной эпителий. Напротив, было обнаружено усиление пролиферации клеток.
в эпителии конъюнктивы, что соответствует гиперплазии, наблюдаемой в
Нокаут рецептора BMP или когда Smad7 управляется промотором K14
(Andl et al., 2004;
He et al., 2002). Таким образом, БМП
обычно подавляют разрастание и дифференциацию конъюнктивы
эпителий и определяют судьбу эпителиальных клеток конъюнктивы.
,
2002). Неспособность вызванной K14 сверхэкспрессии noggin вызывать
EOB может быть вызван поздней экспрессией K14 в конъюнктиве, так же как
веко закрывается, не давая мозгу времени для предотвращения
активация рецепторов BMP.EOB у мышей с дефицитом передачи сигналов BMP был
не из-за снижения пролиферации клеток или увеличения гибели клеток в
глазной эпителий. Напротив, было обнаружено усиление пролиферации клеток.
в эпителии конъюнктивы, что соответствует гиперплазии, наблюдаемой в
Нокаут рецептора BMP или когда Smad7 управляется промотором K14
(Andl et al., 2004;
He et al., 2002). Таким образом, БМП
обычно подавляют разрастание и дифференциацию конъюнктивы
эпителий и определяют судьбу эпителиальных клеток конъюнктивы.
Регулируемая FGF передача сигналов BMP необходима для ядерной локализации фосфорилированного c-Jun и транскрипции
Foxc1, и Foxc2 Фосфорилирование и функция c-Jun в эпителии век зависит от
по крайней мере частично, на Smad-независимую передачу сигналов активином β-B
(Zhang et al., 2003). Тем не мение,
в Acvr1; Bmpr1a DCKO , S mad1 / 5 DCKO и Smad4 CKO веки, c-Jun оказались слабее
фосфорилирован, чем у дикого типа, и не может перемещаться в ядра
клетки перидермы.Предыдущие исследования показали, что Smads может связываться с c-Jun, когда он
не связан с ДНК, что c-Jun фосфорилируется после лечения
клетки с TGFβ и комплексы Smad-c-Jun способствуют AP-1-зависимой
транскрипция (Liberati et al.,
1999; Цин и др.,
2000; Verrecchia et al.,
2001; Чжан и др.,
1998). По нашим данным, BMP, действуя через канонические
Путь R-Smad / Smad4, взаимодействует с активином β-B для обеспечения максимального
фосфорилирование c-Jun и ассоциация с комплексом R-Smad / Smad4
опосредует транслокацию фосфорилированного c-Jun в ядро.Нашим
знаний, это первый отчет, предполагающий, что активированные Smads
необходим для ядерной транслокации c-Jun. Требуются дальнейшие исследования
чтобы определить, важна ли эта функция передачи сигналов BMP в других
примеры слияния эпителия, такие как закрытие нервной трубки и вентральной
закрытие глазного бокала, для которого важна правильная функция пути JNK
существенный (Ся и Карин,
2004 г.).
Активность канонического пути Wnt в веке изменяется в разными путями с помощью передачи сигналов FGF и BMP. ( A-C ) Цельная X-галочка окрашивание репортера TOPGAL дикого типа (A), Fgfr2 CKO (B) и Smad4 CKO (C) веки на E15.5. ( D-F ) Фронтальные участки глаз показаны на A-C. (А) А видная полоса активности Wnt присутствует в верхней части (черная стрелка) и более слабая полоса по краям нижнего века (красная стрелка) глаз дикого типа. Wnt активность также присутствует в фолликулах ресниц (наконечник стрелки) и в эпидермисе. волосяные фолликулы.(B) Активность Wnt равномерно увеличивается в верхнем и нижнем веки мышей Fgfr2 CKO (черные стрелки) и присутствует в эпидермальные волосяные фолликулы (наконечник стрелки). (C) Активность Wnt присутствует в эпидермальные волосяные фолликулы (наконечник стрелки) и эктопические участки в верхней и нижнее веко мышей Smad4 CKO (черные стрелки). (D) Фронтальная срезы показывают, что активность Wnt расположена около переднего края конъюнктива век дикого типа (стрелка).(E) В Fgfr2 CKO веки, активность Wnt распространяется на глазные яблоки конъюнктива (стрелки). (F) В Smad4 CKO веки, Wnt активность присутствует в эктопических волосяных фолликулах (стрелка), что соответствует расположение полосы клеток с высокой активностью Wnt у дикого типа конъюнктива век. Масштабные линейки: 100 мкм.
Передача сигналов BMP была необходима для накопления Foxc1 и Foxc2,
факторы транскрипции, которые, как было показано в предыдущих исследованиях, необходимы для век
закрытие (Kidson et al. , 1999;
Куме и др., 1998;
Смит и др., 2000). Foxc1 и
Foxc2 также экспрессируются в периокулярной мезенхиме.
(Gage et al., 2005) и конечности
бутонная мезенхима (Nifuji et al.,
2001), а обработка Bmp4 и Bmp7 увеличивает
транскрипция Foxc2 в мезенхиме зачатка конечностей в органной культуре
(Nifuji et al., 2001). В
функция Foxc1 и Foxc2 при закрытии век не ясна. Однако у него есть
было показано, что Foxc1 и Foxc2 контролируют сомитогенез, регулируя Notch
передача сигналов (Kume et al., 2001).
Факторы транскрипции Foxc также напрямую стимулируют выработку Notch
лиганд Dll4, чтобы активировать накопление Hey2 в эндотелиальных клетках сосудов
(Хаяси и Куме, 2008 г.).В
эксперименты, предпринятые для другого исследования, мы обнаружили, что делеция в
поверхностная эктодерма пресенилинов Psen1 и Psen2 в фенотипе EOB (см. рис. S1 в дополнительном материале). Потому что в
в большинстве тканей для активации Notch требуется активность пресенилина, Foxc1 и
Foxc2 может способствовать передаче сигналов Notch во время закрытия век. Тем не менее
пресенилины также необходимы для активности других трансмембранных
белки. По этой причине функция передачи сигналов Notch при закрытии века
требует дальнейшего изучения.
, 1999;
Куме и др., 1998;
Смит и др., 2000). Foxc1 и
Foxc2 также экспрессируются в периокулярной мезенхиме.
(Gage et al., 2005) и конечности
бутонная мезенхима (Nifuji et al.,
2001), а обработка Bmp4 и Bmp7 увеличивает
транскрипция Foxc2 в мезенхиме зачатка конечностей в органной культуре
(Nifuji et al., 2001). В
функция Foxc1 и Foxc2 при закрытии век не ясна. Однако у него есть
было показано, что Foxc1 и Foxc2 контролируют сомитогенез, регулируя Notch
передача сигналов (Kume et al., 2001).
Факторы транскрипции Foxc также напрямую стимулируют выработку Notch
лиганд Dll4, чтобы активировать накопление Hey2 в эндотелиальных клетках сосудов
(Хаяси и Куме, 2008 г.).В
эксперименты, предпринятые для другого исследования, мы обнаружили, что делеция в
поверхностная эктодерма пресенилинов Psen1 и Psen2 в фенотипе EOB (см. рис. S1 в дополнительном материале). Потому что в
в большинстве тканей для активации Notch требуется активность пресенилина, Foxc1 и
Foxc2 может способствовать передаче сигналов Notch во время закрытия век. Тем не менее
пресенилины также необходимы для активности других трансмембранных
белки. По этой причине функция передачи сигналов Notch при закрытии века
требует дальнейшего изучения.
Влияние передачи сигналов FGF и BMP на Dkk2 и Sfrp1 выражение в веках. ( A-C ) Гибридизация in situ для Dkk2 транскриптов во фронтальных срезах дикого типа (A), Fgfr2 CKO (B) и Smad4 CKO (C) эмбрионы
на E15.5. (A) Dkk2 уровней транскрипта (синий) были сильными на веке
эпидермиса, бульбарной конъюнктивы и стромы роговицы с более слабым окрашиванием в
конъюнктива век, перидерма и мезенхима у дикого типа.Чувственный зонд
элемент управления показан на вставке. (B, C) Нет значительных изменений в Dkk2 Уровень мРНК обнаружен в Fgfr2 CKO (B) и Smad4 CKO (C) эмбрионы на E15. 5. ( D-F ) Иммуноокрашивание
для белка DKK2 во фронтальных срезах глаза дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) эмбрионы
на E15.5. Белок DKK2 у эмбрионов дикого типа и CKO был подобен белку in situ.
результаты гибридизации.( G-I ) Гибридизация in situ для Sfrp1 транскрипты во фронтальных срезах дикого типа (G), Fgfr2 CKO (H) и Smad4 CKO (I) веки.
В веках дикого типа (G) и Smad4 CKO (I), Sfrp1 транскрипты присутствовали в конъюнктиве (звездочки), сетчатке, сетчатке
пигментный эпителий и хрусталик. В эмбрионах Fgfr2 CKO (H), Sfrp1 транскриптов были легко обнаружены в сетчатке, сетчатке
пигментный эпителий и хрусталик с низким уровнем в конъюнктиве и глазных яблоках
эпителий.Масштабные линейки: 100 мкм.
5. ( D-F ) Иммуноокрашивание
для белка DKK2 во фронтальных срезах глаза дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) эмбрионы
на E15.5. Белок DKK2 у эмбрионов дикого типа и CKO был подобен белку in situ.
результаты гибридизации.( G-I ) Гибридизация in situ для Sfrp1 транскрипты во фронтальных срезах дикого типа (G), Fgfr2 CKO (H) и Smad4 CKO (I) веки.
В веках дикого типа (G) и Smad4 CKO (I), Sfrp1 транскрипты присутствовали в конъюнктиве (звездочки), сетчатке, сетчатке
пигментный эпителий и хрусталик. В эмбрионах Fgfr2 CKO (H), Sfrp1 транскриптов были легко обнаружены в сетчатке, сетчатке
пигментный эпителий и хрусталик с низким уровнем в конъюнктиве и глазных яблоках
эпителий.Масштабные линейки: 100 мкм.
BMP не опосредуют все эффекты передачи сигналов FGF в веках разработка
Хотя функции BMP, которые были идентифицированы в этом исследовании, были в зависимости от Fgfr2, BMP не опосредуют все эффекты передачи сигналов FGF в веках разработка. Fgfr2 необходим в глазной эктодерме для подавления Передача сигналов Wnt, по крайней мере частично, путем индукции антагониста Wnt Sfrp1. Утрата Передача сигналов BMP не влияет на экспрессию Sfrp1 .Эта рука сигнальный путь FGF может включать Shh, поскольку экспрессия Shh в веко модулируется Fgf10 (Tao et al., 2005), и экспрессия Ptch2 , рецептора Shh это прямая цель передачи сигналов Shh, значительно уменьшилась в крышке эктодерма и нижележащая мезенхима век, лишенная Fgfr2 в эктодерма.
Рис. 9. Аномальная дифференцировка эпителия век в Fgfr2 CKO и Smad4 CKO мышей. ( A-C ) Иммуноокрашивание на кератин 14 (K14) во фронтальных срезах от
дикого типа (A), Fgfr2 CKO (B) и Smad4 CKO (C) эмбрионы на E15.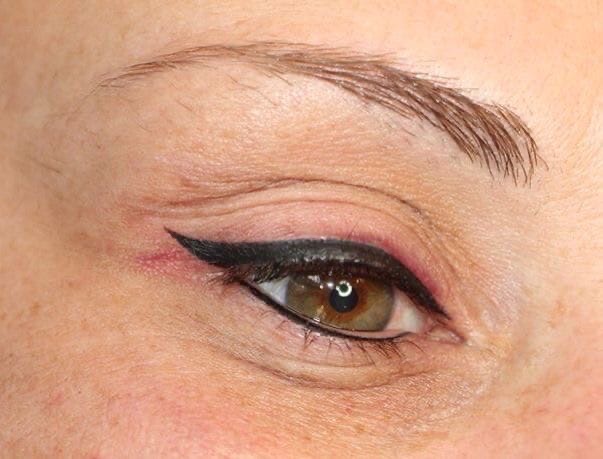 0. В веках дикого типа (A),
Окрашивание K14 было слабым в конъюнктиве век и не обнаруживалось в конъюнктиве век.
бульбарная конъюнктива (стрелки). В Fgfr2 CKO (B) и Smad4 CKO (C) веки, окрашивание K14 было сильным и равномерным
в глазной и бульбарной конъюнктиве (стрелки).( D-F ) Иммуноокрашивание
для кератина 10 (K10) во фронтальных срезах дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) эмбрионы
на E17.5. В веках дикого типа (D) окрашивание K10 наблюдается только в
эпидермис; в конъюнктиве он отрицательный (стрелка). В Fgfr2 CKO (E), K10 обнаружен в эпидермисе и некоторых клетках
глазная конъюнктива, хотя большинство конъюнктивальных клеток не
экспресс К10 (стрелка).В Smad4 CKO (F), K10 обнаружен в
эпидермис и в большинстве клеток конъюнктивы (стрелка). ( G-I )
Иммуноокрашивание на кератин 4 (K4) во фронтальных срезах дикого типа (G), Fgfr2 CKO (H) и Smad4 CKO (I) эмбрионы
на E17.5. В веках дикого типа (G) экспрессия K4 непрерывна в
конъюнктива (стрелка). В Fgfr2 CKO (H), K4 встречается в большинстве
клетки конъюнктивы (стрелка).В Smad4 CKO (I) только K4
обнаруживается в нескольких клетках конъюнктивы; большинство конъюнктивальных клеток не экспрессируют
К4 (стрелка). Масштабные линейки: 100 мкм.
0. В веках дикого типа (A),
Окрашивание K14 было слабым в конъюнктиве век и не обнаруживалось в конъюнктиве век.
бульбарная конъюнктива (стрелки). В Fgfr2 CKO (B) и Smad4 CKO (C) веки, окрашивание K14 было сильным и равномерным
в глазной и бульбарной конъюнктиве (стрелки).( D-F ) Иммуноокрашивание
для кератина 10 (K10) во фронтальных срезах дикого типа (D), Fgfr2 CKO (E) и Smad4 CKO (F) эмбрионы
на E17.5. В веках дикого типа (D) окрашивание K10 наблюдается только в
эпидермис; в конъюнктиве он отрицательный (стрелка). В Fgfr2 CKO (E), K10 обнаружен в эпидермисе и некоторых клетках
глазная конъюнктива, хотя большинство конъюнктивальных клеток не
экспресс К10 (стрелка).В Smad4 CKO (F), K10 обнаружен в
эпидермис и в большинстве клеток конъюнктивы (стрелка). ( G-I )
Иммуноокрашивание на кератин 4 (K4) во фронтальных срезах дикого типа (G), Fgfr2 CKO (H) и Smad4 CKO (I) эмбрионы
на E17.5. В веках дикого типа (G) экспрессия K4 непрерывна в
конъюнктива (стрелка). В Fgfr2 CKO (H), K4 встречается в большинстве
клетки конъюнктивы (стрелка).В Smad4 CKO (I) только K4
обнаруживается в нескольких клетках конъюнктивы; большинство конъюнктивальных клеток не экспрессируют
К4 (стрелка). Масштабные линейки: 100 мкм.
Wnt должна подавляться в глазном эпителии, чтобы
закрытие века (Gage et al.,
2008) (это исследование). Однако полоса сигнальной активности Wnt
обычно присутствует в конъюнктиве глазного яблока у края верхнего
и нижние веки. Эти клетки, по-видимому, включают предшественников
Мейбомиевые железы.Передача сигналов BMP необходима для поддержания этого домена Wnt.
передача сигналов и судьба клеток мейбомиевых желез
(Plikus et al., 2004) (это
изучать). Таким образом, передача сигналов FGF необходима для подавления передачи сигналов Wnt и стимулирования
Передача сигналов BMP, но BMP поддерживают локальную передачу сигналов Wnt в мейбомиевой железе
клетки-предшественники. Факторы, определяющие местоположение и размер BMP и
Передача сигналов Wnt в формировании мейбомиевых желез еще предстоит изучить.
Факторы, определяющие местоположение и размер BMP и
Передача сигналов Wnt в формировании мейбомиевых желез еще предстоит изучить.
Мыши, лишенные передачи сигналов BMP, представляют собой модель заболевания человека. дистихиаз
У мышей, дефицитных по передаче сигналов BMP, эпителиальные клетки конъюнктивы в обоих веки образовывали дополнительный ряд ресниц, характеристика, называемая дистихиаз.Этот фенотип аналогичен фенотипу мышей, у которых избыточная экспрессия ноггин в эктодерме, при которой эктопические ресницы образуются за счет мейбомиевых желез (Plikus et al., 2004 г.). Синдром дистихиаза человека характеризуется: наличие аберрантного второго ряда ресниц на месте мейбомиевых железы (Fox, 1962). Как как следствие, у пациентов наблюдается дисфункция мейбомиевых желез, раздражение роговицы, конъюнктивит и светобоязнь. Большинство семей с дистихиазом имеют общую лимфедему или синдром лимфедема-дистихиаза (ЛД) (OMIM 153400).LD-синдром — аутосомно-доминантное заболевание, вызванное мутациями в FOXC2. Foxc2 гетерозиготных мышей, имитирующих синдром LD, демонстрируя дистихиаз и гиперплазия лимфатических сосудов и лимфатических узлов (Криедерман и др., 2003). В дистихиаз, наблюдаемый в наших исследованиях, согласуется с зависимостью Foxc2 экспрессия в передаче сигналов BMP.
Dkk2 -нулевые мыши также развивают дистихиаз с пониженным Foxc2 выражение в веках
(Гейдж и др., 2008). Экспрессия Dkk2 стимулируется фактором транскрипции Pitx2, который
функционирует в мезенхиме века, происходящей из нервного гребня. Удивительно, но мы
обнаружил мРНК Dkk2 и белок в эпителии век и в
мезенхима. Это наблюдение предполагает, что другой путь регулирует Экспрессия Dkk2 в эктодерме. Избыточная сигнализация Wnt,
в результате дефектов в пути Pitx2 → Dkk2
(Gage et al., 2008), или
Путь Fgf10 → Fgfr2 → Sfrp1, по-видимому, достаточен для подавления Foxc2 , предотвращает закрытие век и вызывает дистихиаз. Поскольку потеря передачи сигналов BMP также приводит к дистихиазу, это кажется возможным.
что чрезмерная активность пути Wnt ингибирует экспрессию Bmp4 в
мезенхима века или функция пути BMP в конъюнктиве
эпителий. Эти возможности еще предстоит проверить.
Поскольку потеря передачи сигналов BMP также приводит к дистихиазу, это кажется возможным.
что чрезмерная активность пути Wnt ингибирует экспрессию Bmp4 в
мезенхима века или функция пути BMP в конъюнктиве
эпителий. Эти возможности еще предстоит проверить.
Передача сигналов BMP необходима для нормальных эпителиальных клеток конъюнктивы судьба
Различная временная и пространственная экспрессия кератиновых промежуточных филаментов является важным аспектом дифференциации и функции многих эпителиев. (Курпакус и др., 1994). В веки дикого типа, эпидермис век выражает K14 перед веком закрытие, тогда как эпителий конъюнктивы начинает экспрессировать K14 как веки закрываются. Однако у мышей, дефицитных по передаче сигналов BMP, конъюнктивальный эпителий экспрессировал K14 одновременно с эпидермисом. Кроме того, клетки конъюнктивы экспрессировали K10, который обычно ограничен эпидермиса, и экспрессия K4 была снижена. Вместе с трансдифференцировка клеток-предшественников мейбомиевых желез в волосяные фолликулы, эти наблюдения предполагают, что передача сигналов BMP обычно предотвращает конъюнктивальный клетки от принятия судьбы эпидермальных клеток (Рис.10А).
Перекрестные помехи между передачей сигналов FGF и BMP могут быть опосредованы Тсс
Предыдущие исследования и результаты, описанные здесь, выявляют сложные
эпителиально-мезенхимальные взаимодействия в развитии век. Активация Fgfr2
в поверхностной эктодерме с помощью Fgf10 из подлежащей мезенхимы
(Tao et al., 2005) требуется
для локальной экспрессии Bmp4 в мезенхиме глазного дна. В
сигнал от эпителия, который способствует экспрессии Bmp4 в
мезенхима, вероятно, будет Shh, так как Shh экспрессируется в веке
наценка от Е13.5 и активная сигнализация Shh (обозначается Ptch2 выражение) имеет паттерн экспрессии, аналогичный паттерну экспрессии Bmp4 в
веко дикого типа. Более того, паттерн остаточной передачи сигналов Shh в Fgfr2 CKO веко аналогично узору остаточного Bmp4 выражение. Эти наблюдения вместе с тем, что
предотвращение передачи сигналов BMP не изменяет функцию Shh, предполагает, что Shh
опосредует Fgfr2-зависимый перекрестный обмен между эпителием и мезенхимой.
Более того, паттерн остаточной передачи сигналов Shh в Fgfr2 CKO веко аналогично узору остаточного Bmp4 выражение. Эти наблюдения вместе с тем, что
предотвращение передачи сигналов BMP не изменяет функцию Shh, предполагает, что Shh
опосредует Fgfr2-зависимый перекрестный обмен между эпителием и мезенхимой.
Резюме роли передачи сигналов BMP и его взаимодействия с другими сигнальные пути, известные или предполагаемые для функционирования в веках мыши разработка. ( A ) В веках с дефицитом передачи сигналов BMP, эпителиальные клетки конъюнктивы экспрессируют эпидермис-специфический кератин 10 и формируют волосяные фолликулы около этого края века. ( B ) Сеть известных или предложены сигнальные пути, контролирующие развитие век.Сделанные наблюдения в этом исследовании отмечены синими стрелками, предыдущие результаты — оранжевыми стрелками. и результаты этого исследования подтверждены зелеными стрелками. Наша модель предполагает, что судьба эпидермальных клеток — это путь по умолчанию, и требуется передача сигналов BMP для предполагаемых эпителиальных клеток конъюнктивы для подавления эпидермального пути дифференцировки и становятся клетками конъюнктивы. Сигнализация BMP не требуется, чтобы инициировать миграцию клеток перидермы по краям века, но требуется для распространения этих клеток по поверхности роговицы.Во время этого процесса передача сигналов BMP необходима для экспрессии Foxc1 и Foxc2, а для полной активации (фосфорилирования) c-Jun. BMP-зависимый образование активных комплексов R-Smad-Smad4 необходимо для транслокации p-c-Jun в ядра клеток перидермы, где, как сообщалось, увеличить экспрессию рецептора эпидермального фактора роста.
Bmp4 сигнализирует вышележащей эктодерме, как показано
фосфорилирование BMP-зависимых R-Smads в эпителии век.Фосфорилированные R-Smads также обнаруживаются в мезенхиме, что позволяет предположить, что BMP
может повлиять на дополнительные цели там. На дополнительном уровне сложности
Паттерны экспрессии Bmp4 и Ptch2 и активность Wnt
разная величина и расположение в верхнем и нижнем веке, и
Экспрессия Bmp4 и Ptch2 была по-разному затронута в
верхнее и нижнее веко удалением Fgfr2 .
На дополнительном уровне сложности
Паттерны экспрессии Bmp4 и Ptch2 и активность Wnt
разная величина и расположение в верхнем и нижнем веке, и
Экспрессия Bmp4 и Ptch2 была по-разному затронута в
верхнее и нижнее веко удалением Fgfr2 .
Сигнальные пути, участвующие в закрытии век
В дополнение к изученным функциям путей FGF, BMP, Shh и Wnt в этом исследовании делеция зародышевой линии показала, что активин β-B, TGFα, HB-EGF и рецептор EGF необходимы для закрытия век. (Luetteke et al., 1994; Luetteke et al., 1993; Миеттинен и др., 1995; Vassalli et al., 1994). Notch передача сигналов также может быть задействована (см. рис. S1 в дополнительном материале). Пути, активируемые этими морфогенами, взаимодействуют очень сложным образом. сети, чтобы обеспечить надлежащую миграцию и слияние небольшой популяции клетки перидермы (рис. 10Б). Необходимы дальнейшие исследования, чтобы полностью определить функции и взаимодействие эти пути. Такие исследования дадут более полное представление о слияние век и, возможно, другие морфогенетические события, зависящие от слияние эпителия, такое как закрытие нервной трубки, зрительной щели, пузырек хрусталика и небные полки.
Сноски
Дополнительные материалы
Дополнительные материалы доступны на сайте http://dev.biologies.org/cgi/content/full/136/10/1741/DC1
Авторы признательны докторам Чжен Махони и Джеффу Майнеру за щедрые обмен реагентами, за многие предложения по техническим аспектам этой работы и за помощь в редактировании. Белинда МакМахан и Джин Джонс подготовили гистологические срезы, а д-р Клаудиа Гарсия предоставила рекомендации по Fgfr2 условных нокаутов, щедро предоставленных доктором Дэвид Орниц, Вашингтонский университет, Сент-Луис. Psen1 и Psen2 флоксированных и мутантных мышей были щедро предоставлены доктором Дж.
Шеном,
Бригам и женская больница, Бостон, Массачусетс, и доктор Рафаэль Копан, Вашингтон
Университет, Сент-Луис. Доктора Питер Грусс и Рут Эшери-Падан создали и
предоставили мышей Le-Cre. Исследование было поддержано
Грант NIH
EY04853 (округ Колумбия) и
Основной грант NIH
P30 EY02687 и неограниченный грант от
Исследования по предотвращению слепоты в отделении
Офтальмология и визуальные науки. Депонировано в ЧВК на
выпуск через 12 месяцев.
 Шеном,
Бригам и женская больница, Бостон, Массачусетс, и доктор Рафаэль Копан, Вашингтон
Университет, Сент-Луис. Доктора Питер Грусс и Рут Эшери-Падан создали и
предоставили мышей Le-Cre. Исследование было поддержано
Грант NIH
EY04853 (округ Колумбия) и
Основной грант NIH
P30 EY02687 и неограниченный грант от
Исследования по предотвращению слепоты в отделении
Офтальмология и визуальные науки. Депонировано в ЧВК на
выпуск через 12 месяцев.
Шеном,
Бригам и женская больница, Бостон, Массачусетс, и доктор Рафаэль Копан, Вашингтон
Университет, Сент-Луис. Доктора Питер Грусс и Рут Эшери-Падан создали и
предоставили мышей Le-Cre. Исследование было поддержано
Грант NIH
EY04853 (округ Колумбия) и
Основной грант NIH
P30 EY02687 и неограниченный грант от
Исследования по предотвращению слепоты в отделении
Офтальмология и визуальные науки. Депонировано в ЧВК на
выпуск через 12 месяцев.Каталожные номера
- ↵
Андл, Т., Ан, К., Кайро, А., Чу, Э.Ю., Вайн-Ли, Л., Редди, С. Т., Крофт, Н. Дж., Себра-Томас, Дж. А., Мецгер, Д., Шамбон, П. и др. (2004). Эпителиальный Bmpr1a регулирует дифференцировку и пролиферация в послеродовых волосяных фолликулах и важна для зуба разработка. Разработка 131,2257 -2268.
- ↵
Эшери-Падан, Р., Марквардт, Т., Чжоу, X. и Gruss, P. (2000). Для линзы требуется активность Pax6 в зачатке хрусталика. формирования и для правильного размещения одиночной сетчатки в глазу. Genes Dev. 14,2701 -2711.
- ↵
Хитил, А., Магнусон, М. А., Райт, К. В. и Моисей, Х. Л. (2002). Условная инактивация рецептора TGF-бета типа II используя Cre: Lox. Бытие 32, 73-75.
- ↵
ДасГупта, Р. и Фукс, Э. (1999). Несколько роли активированных комплексов транскрипции LEF / TCF во время волосяного фолликула развитие и дифференциация. Разработка 126,4557 -4568.
- ↵
Дудас, М., Шридуронгрит, С., Надь, А., Окадзаки, К. и Каартинен, В. (2004). Черепно-лицевые дефекты у мышей отсутствуют. Рецептор BMP типа I Alk2 в клетках нервного гребня. Мех. Dev. 121 173 -182.
- ↵
Fang, J., Dagenais, S.Л., Эриксон, Р. П., Арльт, М. Ф., Глинн, М.
В., Горски, Дж. Л., Сивер, Л. Х. и Гловер, Т. В. (2000).
Мутации в FOXC2 (MFH-1), транскрипционном факторе семейства вилкоголовых, являются
ответственный за наследственный синдром лимфедема-дистихиаз.
Являюсь. J. Hum. Genet. 67 1382
-1388. - ↵
Финдлейтер, Г. С., Макдугалл, Р. Д. и Кауфман, М. Х. (1993). Развитие век, срастание и последующее повторное открытие в мышь. J. Anat. 183 121 -129.
- ↵
Фокс, С. А. (1962). Дистихиаз. Являюсь. J. Ophthalmol. 53,14 -18.
- ↵
Гейдж, П. Дж., Роудс, В., Прука, С. К. и Хьялт, Т. (2005). Карты судьбы нервного гребня и мезодермы у млекопитающих глаз. Вкладывать деньги. Офтальмол. Vis. Sci. 46 4200 -4208.
- ↵
Гейдж, П. Дж., Цянь, М., Ву, Д. и Розенберг, К. И. (2008).Канонический антагонист передачи сигналов Wnt, DKK2, представляет собой существенный эффектор функции PITX2 во время нормального развития глаз. Dev. Биол. 317 310 -324.
- ↵
Гарсия, К. М., Ю, К., Чжао, Х., Ашери-Падан, Р., Орниц, Д. М., Робинсон, М. Л. и Биби, Д. С. (2005). Сигнализация через рецептор FGF-2 требуется для выживания клеток хрусталика и для вывода из клеточного цикла во время дифференцировки клеток волокна хрусталика. Dev. Дин. 233 516 -527.
- ↵
Гауссин, В., Ван де Путте, Т., Мишина, Ю., Хэнкс, М.С., Zwijsen, A., Huylebroeck, D., Behringer, R.R. и Schneider, M.D. (2002). Эндокардиальная подушка и дефекты миокарда после кардиологического миоцит-специфическая условная делеция костного морфогенетического белка рецептор ALK3. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 99 2878 -2883.
- ↵
Хаяси, Х.
и Куме, Т. (2008). Foxc
факторы транскрипции напрямую регулируют экспрессию Dll4 и Hey2 посредством
взаимодействует с сигнальными путями VEGF-Notch в эндотелиальных клетках.PLoS ONE 3, e2401
. - ↵
Хэ, Дж., Шэн, Т., Стелтер, А. А., Ли, К., Чжан, X., Синха, М., Люксон Б. А. и Се Дж. (2006). Подавление передачи сигналов Wnt с помощью hedgehog путь через sFRP-1. J. Biol. Chem. 281,35598 -35602.
- ↵
Хэ, В., Ли, А. Г., Ван, Д., Хань, С., Чжэн, Б., Гуманс, М. Дж., Тен Дийке, П. и Ван, X. J. (2002). Сверхэкспрессия Smad7 приводит к серьезным патологическим изменениям во множестве эпителиальных тканей.EMBO J. 21,2580 -2590.
- ↵
Хуанг, С., Тан, Б., Усоскин, Д., Лехлейдер, Р. Дж., Джамин, С. П., Ли, К., Анзано, М. А., Эбендал, Т., Дэн, К. и Робертс, А. Б. (2002). Условный нокаут гена Smad1. Бытие 32,76 -79.
- ↵
Като, Ю. и Като, М. (2006). Антагонист WNT, SFRP1 является мишенью для передачи сигналов Hedgehog. Int. J. Mol. Med. 17 171 -175.
- ↵
Кидсон, С.Х., Куме, Т., Дэн, К., Уинфри, В. и Хоган, Б. Л. (1999). Ген вилки / крылатой спирали, Mf1, необходим. для нормального развития роговицы и формирования передней камеры в глазе мыши. Dev. Биол. 211 306 -322.
- ↵
Кридерман, Б. М., Майлойд, Т. Л., Витте, М. Х., Дагене, С. Л., Витте, К. Л., Реннельс, М., Бернас, М. Дж., Линч, М. Т., Эриксон, Р. П., Колдер, М.С. и др. (2003). FOXC2 гаплонедостаточные мыши являются моделью синдрома аутосомно-доминантного лимфедема-дистихиаза у человека.Гм. Мол. Genet. 12,1179 -1185.
- ↵
Куме, Т., Дэн, К. Ю., Уинфри, В., Гулд, Д. Б., Уолтер, М.
А.
и Хоган Б. Л. (1998). Ген вилкоголовой / крылатой спирали Mf1
Нарушается мутация плейотропной мыши врожденной гидроцефалией.
Ячейка 93,985
-996. - ↵
Куме, Т., Цзян, Х., Топчевска, Дж. М. и Хоган, Б. Л. (2001). Факторы транскрипции крылатой спирали мыши, Foxc1 и Foxc2 необходимы для развития сердечно-сосудистой системы и сомитогенеза.Genes Dev. 15 2470 -2482.
- ↵
Курпакус, М. А., Маниачи, М. Т. и Эско, М. (1994). Экспрессия кератинов K12, K4 и K14 во время развития эпителия глазной поверхности. Curr. Eye Res. 13 805 -814.
- ↵
Li, C., Guo, H., Xu, X., Weinberg, W. и Deng, C. X. (2001). Рецептор 2 фактора роста фибробластов (Fgfr2) играет роль играет важную роль в формировании и формировании рисунка век и кожи.Dev. Дин. 222 471 -483.
- ↵
Ли, Г., Густафсон-Браун, К., Хэнкс, С. К., Насон, К., Арбейт, Дж. М., Польяно, К., Уиздом, Р. М. и Джонсон, Р. С. (2003). c-Jun необходим для организации переднего края эпидермиса. Dev. Ячейка 4,865 -877.
- ↵
Либерати, Н. Т., Датто, М. Б., Фредерик, Дж. П., Шен, X., Вонг, К., Ружье-Чепмен, Э. М. и Ван, Х. Ф. (1999). Smads bind непосредственно к семейству Jun факторов транскрипции AP-1.Proc. Natl. Акад. Sci. США 96,4844 -4849.
- ↵
Липински, Р. Дж., Хатсон, П. Р., Ханнам, П. В., Нидза, Р. Дж., Вашингтон, И. М., Мур, Р. В., Гирдаукас, Г. Г., Петерсон, Р. Э. и Бушман, W. (2008). Тератогенность, зависящая от дозы и пути введения, токсичность и фармакокинетические профили сигнального антагониста hedgehog циклопамин у мышей. Toxicol. Sci. 104 189 -197.
- ↵
Люетеке, Н.К., Цю, Т.
Х., Пайффер, Р. Л., Оливер, П.,
Смитис, О. и Ли, Д. С. (1993). Дефицит TGF альфа
приводит к аномалиям волосяного фолликула и глаз у мышей-мишеней и мышей-1.
Ячейка 73,263
-278. - ↵
Люттеке, Н. К., Филлипс, Х. К., Цю, Т. Х., Коупленд, Н. Г., Эрп, Х.С., Дженкинс, Н.А. и Ли, Д.С. (1994). Мышь Фенотип waved-2 является результатом точечной мутации тирозина рецептора EGF киназа. Genes Dev. 8,399 -413.
- ↵
Миеттинен, П. Дж., Бергер, Дж. Э., Менесес, Дж., Фунг, Ю., Педерсен Р. А., Верб З. и Деринк Р. (1995). Эпителиальный незрелость и полиорганная недостаточность у мышей, лишенных эпидермального фактора роста рецептор. Природа 376 337 -341.
- ↵
Майн, Н., Ивамото, Р. и Мекада, Е. (2005). HB-EGF способствует миграции эпителиальных клеток в развитии век. Развитие 132,4317 -4326.
- ↵
Нифудзи, А., Миура, Н., Като, Н., Келлерманн, О. и Нода, М. (2001). Костная регуляция морфогенетических белков вилки / крылатого фактор транскрипции спирали Foxc2 (Mfh2) в мышиной мезодермальной клеточной линии C1 и в клетках-предшественниках скелета. J. Bone Miner. Res. 16,1765 -1771.
- ↵
Пик, Э., Джу, В. Дж., Хейер, Дж., Эскаланте-Алькальд, Д., Стюарт, К. Л., Вайнштейн, М., Дэн, К., Кучерлапати, Р., Боттингер, Э. П. и Робертс, А. Б. (2001). Функциональная характеристика трансформирует передачу сигналов фактора роста бета у Smad2- и Smad3-дефицитных фибробласты. J. Biol. Chem. 276,19945 -19953.
- ↵
Пликус, М., Ван, В. П., Лю, Дж., Ван, X., Цзян, Т.-Х. и Чуонг, Ч.-М. (2004). Морфорегуляция эктодермальных органов: патология покровов и фенотипические вариации у мышей, сконструированных с помощью K14-noggin через модуляцию костного морфогенного белкового пути.
Являюсь. Дж.
Патол. 164 1099
-1114. - ↵
Цин, Дж., Чжан, Ю. и Деринк, Р. (2000). Структурно-функциональная характеристика трансформирующего роста Фактор-бета-индуцированная транскрипционная кооперативность Smad3 / c-Jun. Дж. Биол. Chem. 275,38802 -38812.
- ↵
Робертс, А. Б., Тиан, Ф., Байфилд, С. Д., Стуэлтен, К., Оошима, А., Сайка С. и Фландерс К. С. (2006). Smad3 является ключом к TGF-бета-опосредованный переход эпителия в мезенхиму, фиброз, опухоль подавление и метастазирование.Фактор роста цитокинов Ред. 17,19 -27.
- ↵
Шаров А.А., Вайнер Л., Шарова Т.Ю., Зибенхаар Ф., Атоян, Р., Регинато, А. М., Макнамара, К. А., Фуна, К., Гилкрест, Б. А., Brissette, J. L. et al. (2003). Сверхэкспрессия Noggin препятствует открытию век за счет изменения апоптоза и дифференцировки эпидермиса. EMBO J. 22,2992 -3003.
- ↵
Смит, Р.С., Забалета, А., Куме, Т., Савинова, О.В., Кидсон, С. Х., Мартин, Дж. Э., Нисимура, Д. Ю., Алвард, В. Л., Хоган, Б. Л. и Джон, S. W. (2000). Гаплонедостаточность факторов транскрипции FOXC1 и FOXC2 приводят к аберрантному развитию глаз. Гм. Мол. Genet. 9,1021 -1032.
- ↵
Штайнер, Х., Дафф, К., Капелл, А., Ромиг, Х., Грим, М. Г., Линкольн С., Харди Дж., Ю X., Пиччиано М., Фехтелер К. и др. (1999). Мутация пресенилина-2 с потерей функции мешает с образованием амилоидного бета-пептида и передачей сигналов notch.J. Biol. Chem. 274,28669 -28673.
- ↵
Суинделл, Э. К., Лю, К., Шах, Р., Смит, А. Н., Ланг, Р. А. и Jamrich, M. (2008). Формирование глаз при отсутствии сетчатка. Dev. Биол. 322,56 -64.
- ↵
Такатори, А.
, Гех, Э., Чен, Л., Чжан, Л., Меллер, Дж. И Ся,
Ю. (2008). Дифференциальная передача MEKK1 морфогенетическая
сигналы от JNK1 и JNK2. Разработка
135, 23-32. - ↵
Тао, Х., Симидзу, М., Кусумото, Р., Оно, К., Нодзи, С. и Охучи, Х. (2005). Двойная роль FGF10 в пролиферации и скоординированная миграция эпителиальных клеток ведущего края века мыши разработка. Разработка 132,3217 -3230.
- ↵
Umans, L., Vermeire, L., Francis, A., Chang, H., Huylebroeck, D. и Zwijsen, A. (2003). Генерация гибкого аллеля Smad5 для обусловленного Cre-опосредованного нокаута у мыши. Бытие 37,5 -11.
- ↵
Вассалли, А., Мацук, М. М., Гарднер, Х. А., Ли, К. Ф. и Jaenisch, R. (1994). Ген бета-субъединицы B активина / ингибина нарушение ведет к дефектам развития век и женского размножения. Genes Dev. 8 414 -427.
- ↵
Verrecchia, F., Vindevoghel, L., Lechleider, R.J., Uitto, J., Робертс, А. Б. и Мовьель, А. (2001). Smad3 / AP-1 взаимодействия контролируют транскрипционные ответы на TGF-бета в промотор-специфический способ.Онкоген 20,3332 -3340.
- ↵
Уэстон, К. Р., Вонг, А., Холл, Дж. П., Гоуд, М. Э., Флавелл, Р. А. и Дэвис Р. Дж. (2004). C-Jun Nh3-терминальная киназа представляет собой необходим для экспрессии эпидермального фактора роста во время эпидермального морфогенез. Proc. Natl. Акад. Sci. Соединенные Штаты Америки 101,14114 -14119.
- ↵
Ся Ю. и Карин М. (2004). Контроль над подвижность клеток и морфогенез эпителия под действием киназ Jun.Тенденции Cell Biol. 14,94 -101.
- ↵
Янг, Х., Ли, К., Эррера, П. Л. и Дэн, К. Х. (2002).
Генерация мышей с условным нокаутом Smad4 / Dpc4.
Бытие 32,80
-81. - ↵
Ю, Х., Саура, К. А., Чой, С. Ю., Сун, Л. Д., Янг, X., Хэндлер, М., Каварабаяши, Т., Юнкин, Л., Феделес, Б., Уилсон, М.А. и др. al. (2001). Процессинг APP и синаптическая пластичность в мышам с условным нокаутом пресенилин-1.Нейрон 31 713 -726.
- ↵
Ю, К., Сюй, Дж., Лю, З., Сосич, Д., Шао, Дж., Олсон, Э. Н., Таулер, Д. А. и Орниц, Д. М. (2003). Условный инактивация рецептора 2 FGF показывает важную роль передачи сигналов FGF в регуляция функции остеобластов и роста костей. Развитие 130,3063 -3074.
- ↵
Зенц, Р., Шойх, Х., Мартин, П., Франк, К., Эферл, Р., Кеннер, Л., Сибилия, М.и Вагнер, Э. Ф. (2003). c-Jun регулирует закрытие век и развитие опухоли кожи посредством передачи сигналов EGFR. Dev. Ячейка 4,879 -889.
- ↵
Чжан, Л., Ван, В., Хаяши, Ю., Шут, Дж. В., Бирк, Д. Э., Гао, М., Лю, К. Ю., Као, В. В., Карин, М. и Ся, Ю. (2003). Роль MEK-киназы 1 в индуцированном TGF-бета / активином движение эпителия и закрытие век эмбриона. EMBO J. 22,4443 -4454.
- ↵
Чжан, Ю., Фен X. и Деринк Р. (1998). Smad3 и Smad4 взаимодействуют с c-Jun / c-Fos, чтобы опосредовать индуцированный TGF-бета транскрипция. Природа 394 909 -913.
 В., Горски, Дж. Л., Сивер, Л. Х. и Гловер, Т. В. (2000).
Мутации в FOXC2 (MFH-1), транскрипционном факторе семейства вилкоголовых, являются
ответственный за наследственный синдром лимфедема-дистихиаз.
Являюсь. J. Hum. Genet. 67 1382
-1388.
В., Горски, Дж. Л., Сивер, Л. Х. и Гловер, Т. В. (2000).
Мутации в FOXC2 (MFH-1), транскрипционном факторе семейства вилкоголовых, являются
ответственный за наследственный синдром лимфедема-дистихиаз.
Являюсь. J. Hum. Genet. 67 1382
-1388.  и Куме, Т. (2008). Foxc
факторы транскрипции напрямую регулируют экспрессию Dll4 и Hey2 посредством
взаимодействует с сигнальными путями VEGF-Notch в эндотелиальных клетках.PLoS ONE 3, e2401
.
и Куме, Т. (2008). Foxc
факторы транскрипции напрямую регулируют экспрессию Dll4 и Hey2 посредством
взаимодействует с сигнальными путями VEGF-Notch в эндотелиальных клетках.PLoS ONE 3, e2401
.  Х., Пайффер, Р. Л., Оливер, П.,
Смитис, О. и Ли, Д. С. (1993). Дефицит TGF альфа
приводит к аномалиям волосяного фолликула и глаз у мышей-мишеней и мышей-1.
Ячейка 73,263
-278.
Х., Пайффер, Р. Л., Оливер, П.,
Смитис, О. и Ли, Д. С. (1993). Дефицит TGF альфа
приводит к аномалиям волосяного фолликула и глаз у мышей-мишеней и мышей-1.
Ячейка 73,263
-278.  Являюсь. Дж.
Патол. 164 1099
-1114.
Являюсь. Дж.
Патол. 164 1099
-1114.  , Гех, Э., Чен, Л., Чжан, Л., Меллер, Дж. И Ся,
Ю. (2008). Дифференциальная передача MEKK1 морфогенетическая
сигналы от JNK1 и JNK2. Разработка
135, 23-32.
, Гех, Э., Чен, Л., Чжан, Л., Меллер, Дж. И Ся,
Ю. (2008). Дифференциальная передача MEKK1 морфогенетическая
сигналы от JNK1 и JNK2. Разработка
135, 23-32.  Генерация мышей с условным нокаутом Smad4 / Dpc4.
Бытие 32,80
-81.
Генерация мышей с условным нокаутом Smad4 / Dpc4.
Бытие 32,80
-81. Воспаление век — AMBOSS
Последнее обновление: 23 февраля 2021 г.
Резюме
Веки содержат различные железы (мейбомиевы, Зейсовы и Молля), секреты которых служат компонентами слезной пленки и помощь в смазке век. Бактериальные инфекции или накопление / нарушение регуляции секреции могут привести к трем распространенным воспалительным состояниям века: ячмень (ячмень), блефарит и халязион.Гордеолум — это острое воспаление век (обычно инфекционное), связанное с болезненными, эритематозными и заполненными гноем узелками. Блефарит — это хроническое (обычно инфекционное) воспалительное состояние, связанное с твердыми, чешуйчатыми бляшками и / или масляными отложениями на краях век. Халязион — это очаговая стерильная гранулематозная опухоль сальной железы века, которая может быть результатом гордеолума или блефарита. Диагноз обычно ставят клинически. Лечение зависит от конкретного состояния и включает гигиену век, местные антибиотики, если подозревается инфекционная этиология, или хирургическое вмешательство.
Блефарит — это хроническое (обычно инфекционное) воспалительное состояние, связанное с твердыми, чешуйчатыми бляшками и / или масляными отложениями на краях век. Халязион — это очаговая стерильная гранулематозная опухоль сальной железы века, которая может быть результатом гордеолума или блефарита. Диагноз обычно ставят клинически. Лечение зависит от конкретного состояния и включает гигиену век, местные антибиотики, если подозревается инфекционная этиология, или хирургическое вмешательство.
Гордеолум (ячмень)
- Определение: распространенное острое воспаление слезной железы или фолликулов ресниц (Зейса или мейбомиевых желез)
- Этиология
- Классификация
- Клинические особенности
- Диагностика: обычно клинический диагноз
- Лечение: большинство ячменей проходят самостоятельно и проходят спонтанно через 1-2 недели.
- Осложнения
Ссылки: [1] [2]
Блефарит (плоский блефарит)
- Определение: блефарит — это распространенное заболевание век, которое проявляется как хроническое и / или рецидивирующее чешуйчатое воспаление краев век.
- Этиология
- Стафилококки (наиболее частый возбудитель)
- Другая инфекционная этиология: вирусная (например, простой герпес или ветряная оспа) или паразитарная (демодекоз или фтириаз пальпебрарум).
- Аллергические реакции или анатомическая непроходимость сальных желез
- Эндогенные факторы: жирные отложения, себорея, гиперсекреция век век, скопление секрета мейбомиевых желез.
- Внешние факторы: дым, пыль, сухой климат в помещении, химические вещества, токсичность лекарств
- Системные заболевания: розацеа, атопия и себорейный дерматит
- Заболевания глаз: синдромы сухого глаза (например,g., синдром Шегрена), халязион, трихиаз, конъюнктивит и кератит
- Классификация
- Клинические признаки
- Хронические или повторяющиеся красные опухшие веки и неровные края век
- Корки, чешуйчатые бляшки и / или жирные отложения на краях век и ресницах
- Раздражение глаз и нарушения зрения
- Диагностика: клинический диагноз
- Лечение
- Осложнения
Ссылки: [1] [3] [4] [5] [6]
Халазион (липогранулема мейбомиевых желез)
- Определение: очаговая гранулематозная опухоль сальных желез без некроза, которая в первую очередь вызвана закупоркой желез.
- Этиология
- Клинические особенности
- Хронический (медленнорастущий) твердый эластичный узелок на веке
- Тяжесть века
- Может вызвать нарушение зрения, если он достаточно большой
- Диагностика
- Обычно клинический диагноз
- Выворачивание века может улучшить визуализацию поражения.
- Биопсия: для стойкого или рецидивирующего халязиона (может указывать на лежащую в основе карциному века)
- Лечение

Стойкий или рецидивирующий халязион может быть признаком сальной карциномы (карциномы мейбомиевой железы). Халязион может также клинически напоминать базальноклеточную карциному!
Каталожные номера: [1] [7] [8] [9] [10]
Список литературы
- Блефарит. https://www.msdmanuals.com/professional/eye-disorders/eyelid-and-lacrimal-disorders/blepharitis . Обновлено: 1 января 2018 г. Доступ: 13 июня 2018 г.
- Лоури Р.С., Даль А.А. Блефарит у взрослых. Блефарит у взрослых . Нью-Йорк, штат Нью-Йорк: WebMD. http://emedicine.medscape.com/article/1211763 . Обновлено: 15 декабря 2016 г. Дата обращения: 23 мая 2017 г.
- Shtein RM, Trobe J, Libman H. Blepharitis. В: Сообщение TW, под ред. Дата обновления .Уолтем, Массачусетс: UpToDate. http://www.uptodate.com/contents/blepharitis . Последнее обновление: 28 февраля 2017 г. Дата обращения: 23 мая 2017 г.
- Хурана АК. Комплексная офтальмология . Аншань Паб ; 2008 г.
- Руководство по предпочтительной практике лечения блефарита.
